
Biosynthesis of the Novel Endogenous 15-Lipoxygenase Metabolites N-13-Hydroxy-octodecadienoyl-ethanolamine and 13-Hydroxy-octodecadienoyl-glycerol by Human Neutrophils and Eosinophils

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis and Purification of 1-LG-d5
2.3. Synthesis and Purification of 13-HODE-G and 13-HODE-G-d5
2.4. Characterization with 1D and 2D NMR
2.5. Characterization with HR-ESI-MS/MS Spectrometry
2.6. Enzymatic Assays
2.7. Analysis of 13-HODE-G and 13-HODE-EA by LC-MS/MS
2.8. Isolation of Human Neutrophils and Eosinophils
2.9. Cell Stimulations
2.10. CB1 and CB2 Binding Assays
2.11. Analysis of PPAR-α, PPAR-γ and TRPV1 Functional Activity
2.12. Computational Analysis of PPAR-α Binding
2.13. Ethics Committee Approval
3. Results
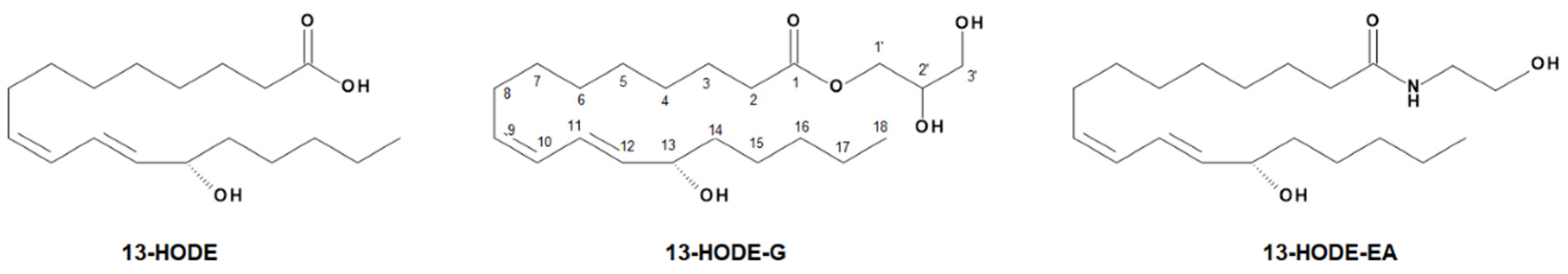
3.1. Synthesis of 1-LG-d5, 13-HODE-G, and 13-HODE-d5
3.2. Biosynthesis of 13-HODE-G by Human Recombinant 15-LO-1 and 15-LO-2
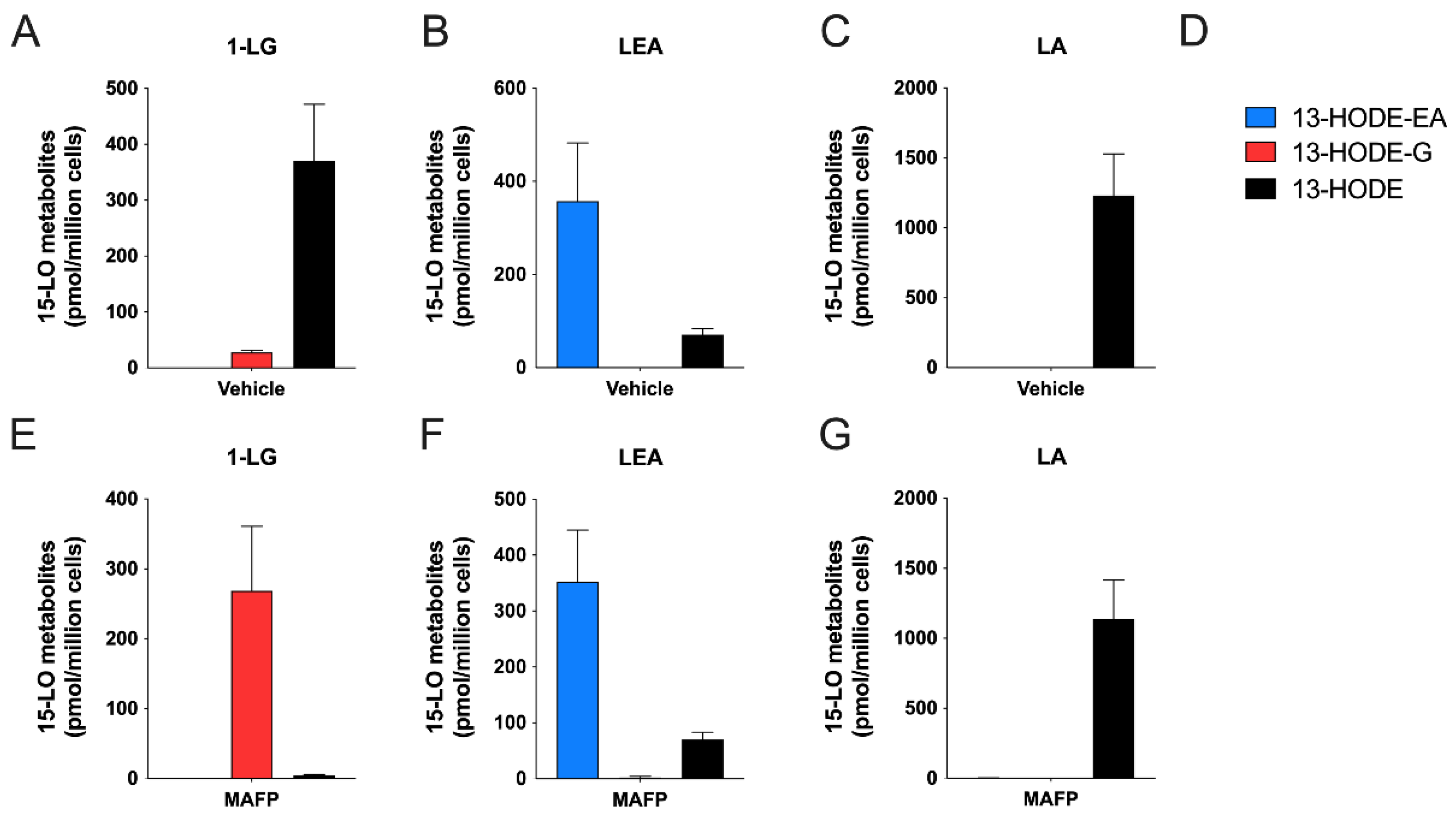
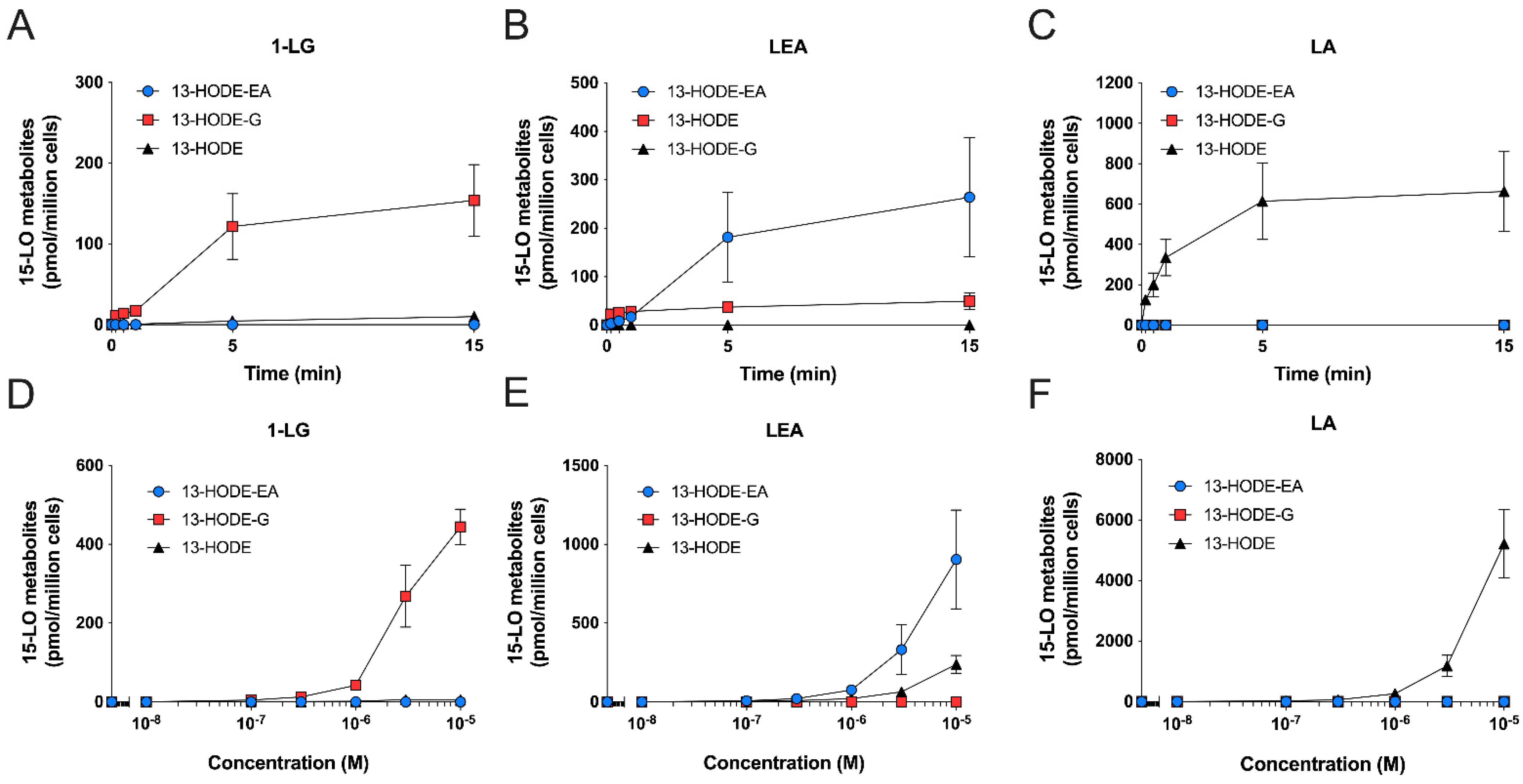
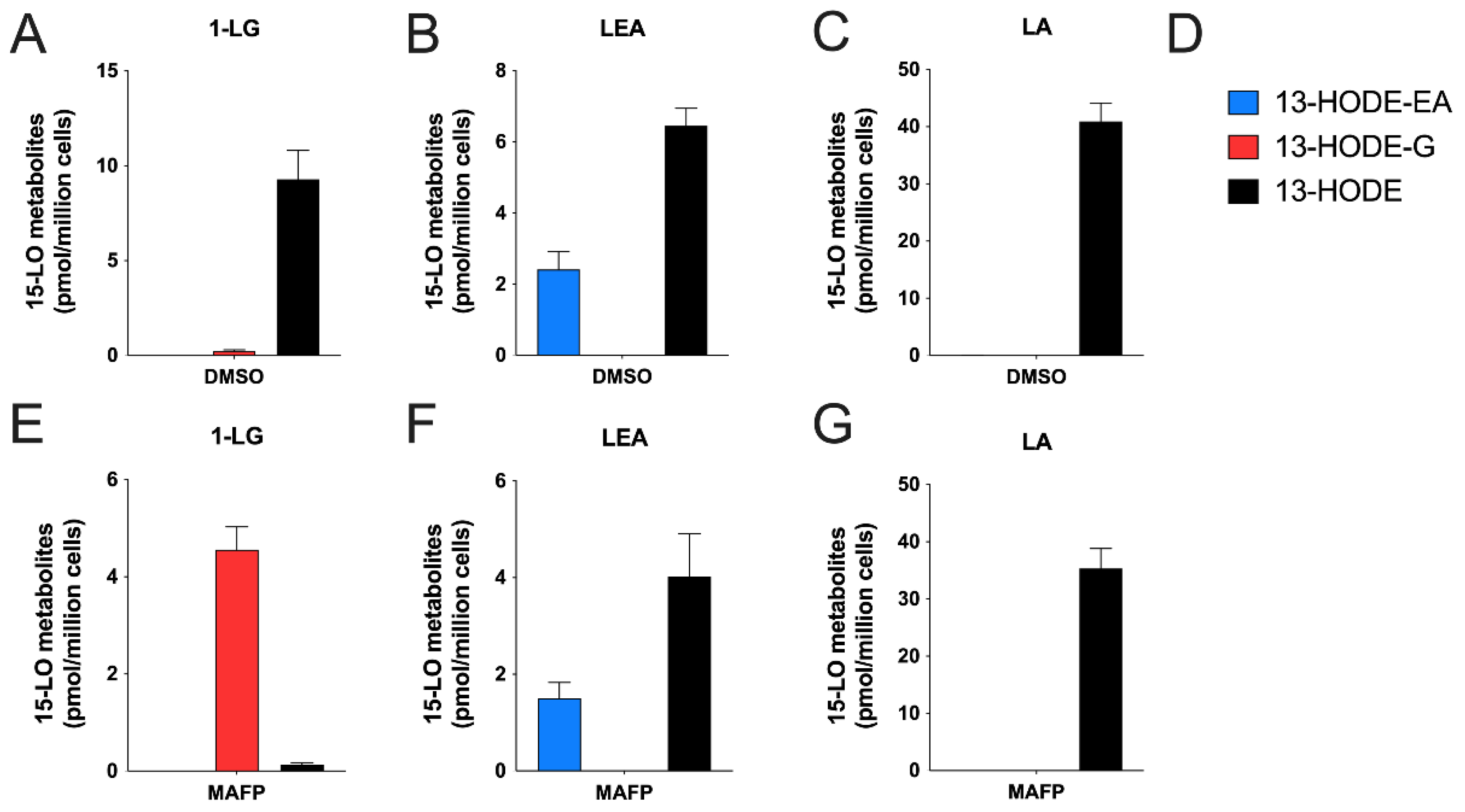
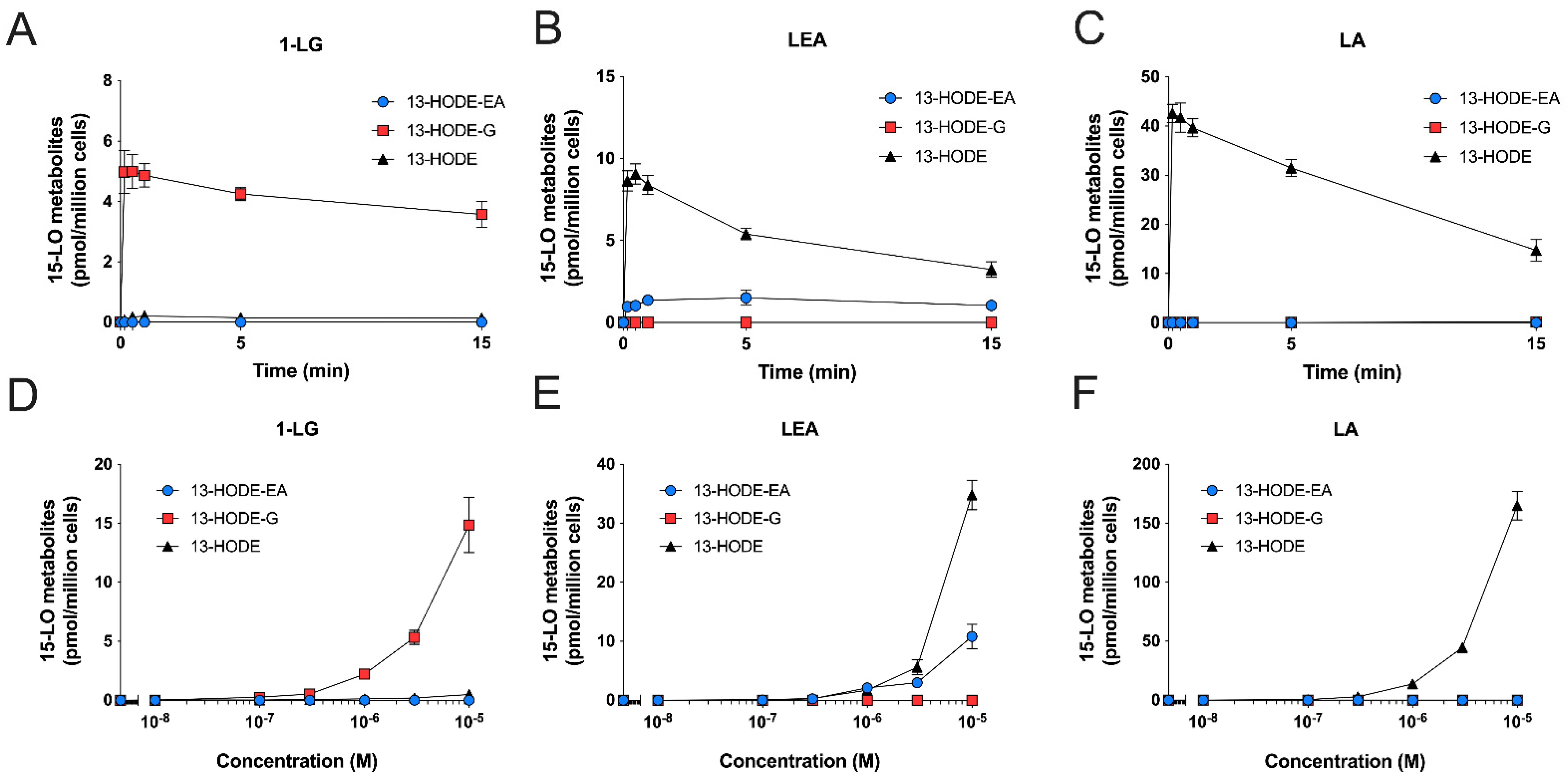
3.3. Human Eosinophils Metabolize LA, 1-LG, and LEA via the 15-LO Pathway
3.4. Human Neutrophils Also Metabolize LA, 1-LG, and LEA into 13-Hydroxylated Compounds
3.5. Substrate Preference of Eosinophils and Neutrophils
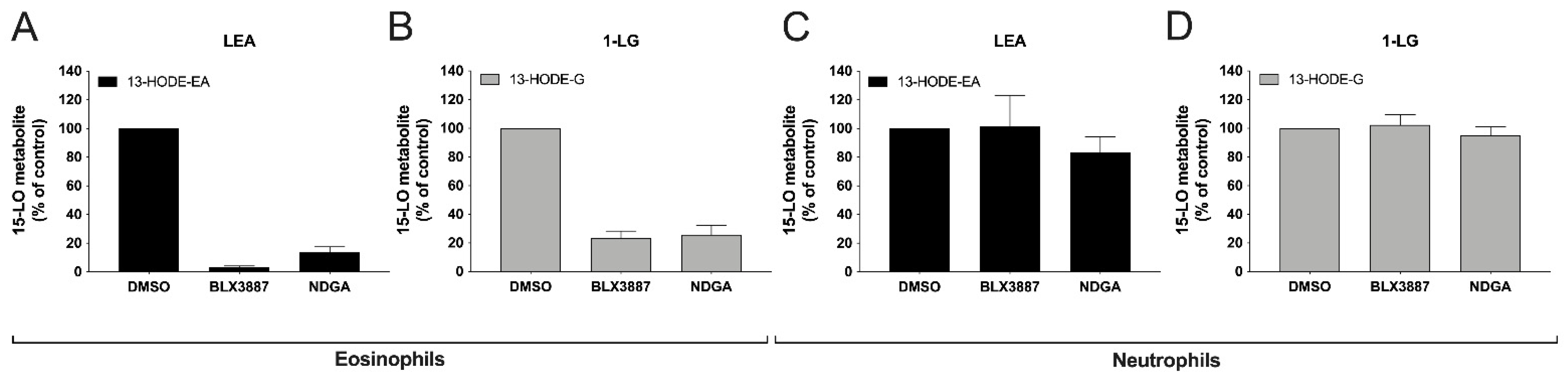
3.6. Inhibition of 13-HODE-EA and 13-HODE-G Biosynthesis by BLX-3887 and NDGA
3.7. Detection of 13-HODE-G In Vivo
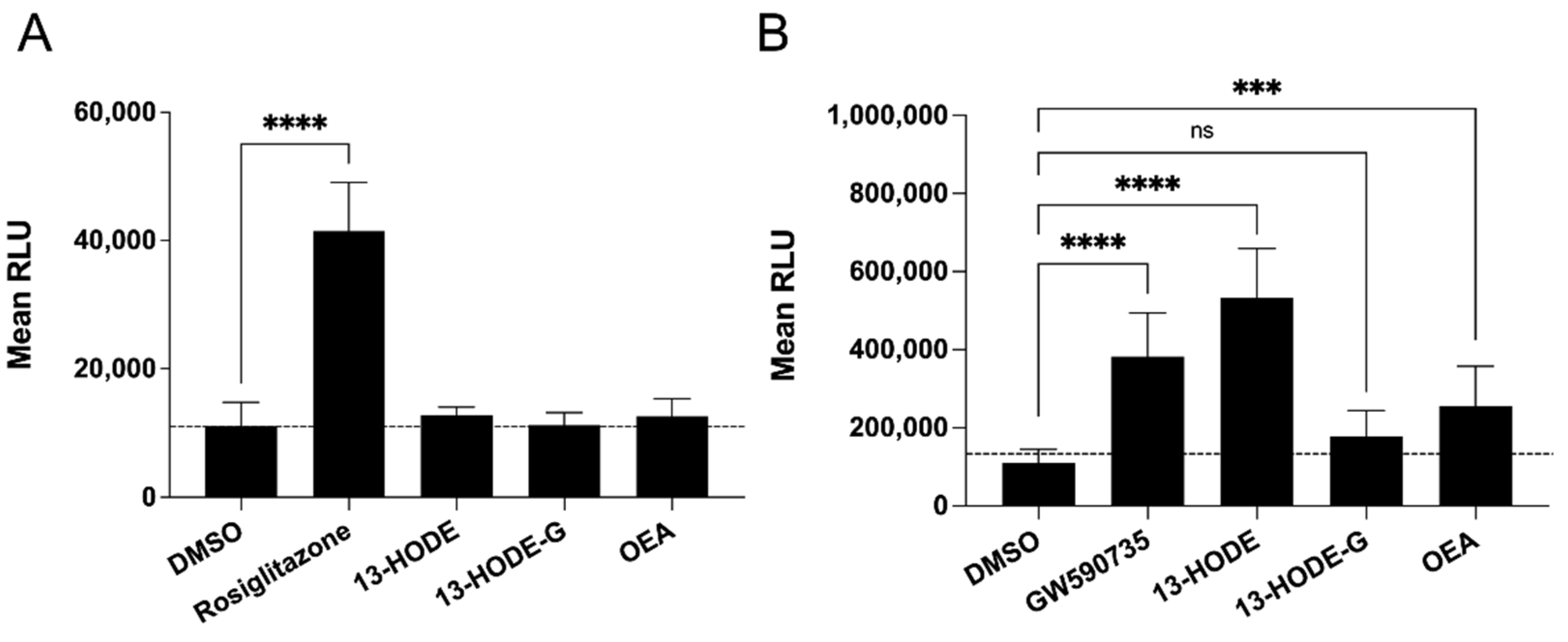
3.8. Binding Assays
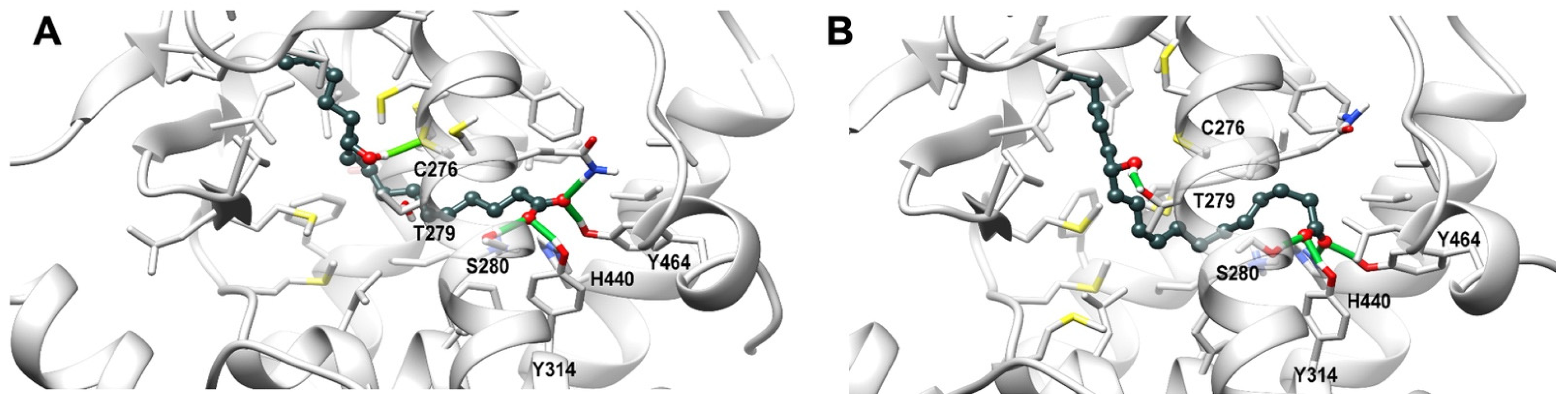
3.9. Theoretical Complex of PPARα with 13-HODE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turcotte, C.; Chouinard, F.; Lefebvre, J.S.; Flamand, N. Regulation of inflammation by cannabinoids, the endocannabinoids 2-arachidonoyl-glycerol and arachidonoyl-ethanolamide, and their metabolites. J. Leukoc. Biol 2015, 97, 1049–1070. [Google Scholar] [CrossRef]
- Di Marzo, V. New approaches and challenges to targeting the endocannabinoid system. Nat. Rev. Drug Discov. 2018, 17, 623–639. [Google Scholar] [CrossRef]
- Prescott, S.M.; Majerus, P.W. Characterization of 1,2-diacylglycerol hydrolysis in human platelets. Demonstration of an arachidonoyl-monoacylglycerol intermediate. J. Biol. Chem. 1983, 258, 764–769. [Google Scholar] [CrossRef]
- Goparaju, S.K.; Ueda, N.; Taniguchi, K.; Yamamoto, S. Enzymes of porcine brain hydrolyzing 2-arachidonoylglycerol, an endogenous ligand of cannabinoid receptors. Biochem. Pharmacol. 1999, 57, 417–423. [Google Scholar] [CrossRef]
- Xie, S.; Borazjani, A.; Hatfield, M.J.; Edwards, C.C.; Potter, P.M.; Ross, M.K. Inactivation of lipid glyceryl ester metabolism in human THP1 monocytes/macrophages by activated organophosphorus insecticides: Role of carboxylesterases 1 and 2. Chem. Res. Toxicol. 2010, 23, 1890–1904. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Borazjani, A.; Matthews, A.T.; Mangum, L.C.; Edelmann, M.J.; Ross, M.K. Identification of palmitoyl protein thioesterase 1 in human THP1 monocytes and macrophages and characterization of unique biochemical activities for this enzyme. Biochemistry 2013, 52, 7559–7574. [Google Scholar] [CrossRef] [Green Version]
- Savinainen, J.R.; Patel, J.Z.; Parkkari, T.; Navia-Paldanius, D.; Marjamaa, J.J.; Laitinen, T.; Nevalainen, T.; Laitinen, J.T. Biochemical and pharmacological characterization of the human lymphocyte antigen B-associated transcript 5 (BAT5/ABHD16A). PLoS ONE 2014, 9, e109869. [Google Scholar] [CrossRef]
- Navia-Paldanius, D.; Savinainen, J.R.; Laitinen, J.T. Biochemical and pharmacological characterization of human alpha/beta-hydrolase domain containing 6 (ABHD6) and 12 (ABHD12). J. Lipid Res. 2012, 53, 2413–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cravatt, B.F.; Giang, D.K.; Mayfield, S.P.; Boger, D.L.; Lerner, R.A.; Gilula, N.B. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996, 384, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.Q.; Mikkelsen, T.S.; McKinney, M.K.; Lander, E.S.; Cravatt, B.F. A second fatty acid amide hydrolase with variable distribution among placental mammals. J. Biol. Chem. 2006, 281, 36569–36578. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, K.; Sun, Y.X.; Okamoto, Y.; Araki, N.; Tonai, T.; Ueda, N. Molecular characterization of N-acylethanolamine-hydrolyzing acid amidase, a novel member of the choloylglycine hydrolase family with structural and functional similarity to acid ceramidase. J. Biol. Chem. 2005, 280, 11082–11092. [Google Scholar] [CrossRef] [Green Version]
- Chouinard, F.; Turcotte, C.; Guan, X.; Larose, M.C.; Poirier, S.; Bouchard, L.; Provost, V.; Flamand, L.; Grandvaux, N.; Flamand, N. 2-Arachidonoyl-glycerol- and arachidonic acid-stimulated neutrophils release antimicrobial effectors against E. coli, S. aureus, HSV-1, and RSV. J. Leukoc. Biol. 2013, 93, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouinard, F.; Lefebvre, J.S.; Navarro, P.; Bouchard, L.; Ferland, C.; Lalancette-Hebert, M.; Marsolais, D.; Laviolette, M.; Flamand, N. The endocannabinoid 2-arachidonoyl-glycerol activates human neutrophils: Critical role of its hydrolysis and de novo leukotriene B4 biosynthesis. J. Immunol. 2011, 186, 3188–3196. [Google Scholar] [CrossRef] [Green Version]
- Larose, M.C.; Turcotte, C.; Chouinard, F.; Ferland, C.; Martin, C.; Provost, V.; Laviolette, M.; Flamand, N. Mechanisms of human eosinophil migration induced by the combination of IL-5 and the endocannabinoid 2-arachidonoyl-glycerol. J. Allergy Clin. Immunol. 2014, 133, 1480–1482. [Google Scholar] [CrossRef]
- Kudalkar, S.N.; Kingsley, P.J.; Marnett, L.J. Assay of Endocannabinoid Oxidation by Cyclooxygenase-2. Methods Mol. Biol. 2016, 1412, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.E.; Kim, J.S.; Das, A. Emerging class of omega-3 fatty acid endocannabinoids & their derivatives. Prostaglandins Other Lipid Mediat. 2019, 143, 106337. [Google Scholar] [CrossRef]
- Moody, J.S.; Kozak, K.R.; Ji, C.; Marnett, L.J. Selective oxygenation of the endocannabinoid 2-arachidonylglycerol by leukocyte-type 12-lipoxygenase. Biochemistry 2001, 40, 861–866. [Google Scholar] [CrossRef]
- Edgemond, W.S.; Hillard, C.J.; Falck, J.R.; Kearn, C.S.; Campbell, W.B. Human platelets and polymorphonuclear leukocytes synthesize oxygenated derivatives of arachidonylethanolamide (anandamide): Their affinities for cannabinoid receptors and pathways of inactivation. Mol. Pharmacol. 1998, 54, 180–188. [Google Scholar] [CrossRef]
- Kozak, K.R.; Gupta, R.A.; Moody, J.S.; Ji, C.; Boeglin, W.E.; DuBois, R.N.; Brash, A.R.; Marnett, L.J. 15-Lipoxygenase metabolism of 2-arachidonylglycerol. Generation of a peroxisome proliferator-activated receptor alpha agonist. J. Biol. Chem. 2002, 277, 23278–23286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archambault, A.S.; Turcotte, C.; Martin, C.; Provost, V.; Larose, M.C.; Laprise, C.; Chakir, J.; Bissonnette, E.; Laviolette, M.; Bosse, Y.; et al. Comparison of eight 15-lipoxygenase (LO) inhibitors on the biosynthesis of 15-LO metabolites by human neutrophils and eosinophils. PLoS ONE 2018, 13, e0202424. [Google Scholar] [CrossRef] [Green Version]
- Tinto, F.; Archambault, A.S.; Dumais, E.; Rakotoarivelo, V.; Kostrzewa, M.; Martin, C.; Plante, P.L.; Desjardins, Y.; Simard, M.; Pouliot, R.; et al. Synthesis and molecular targets of N-13-hydroxy-octadienoyl-ethanolamine, a novel endogenous bioactive 15-lipoxygenase-derived metabolite of N-linoleoyl-ethanolamine found in the skin and saliva. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158954. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Fredman, G.; Krishnamoorthy, S.; Agrawal, N.; Irimia, D.; Piomelli, D.; Serhan, C.N. Decoding functional metabolomics with docosahexaenoyl ethanolamide (DHEA) identifies novel bioactive signals. J. Biol. Chem. 2011, 286, 31532–31541. [Google Scholar] [CrossRef] [Green Version]
- Engels, F.; Kessels, G.C.; Henricks, P.A.; Nijkamp, F.P. Preferential formation of 13-hydroxylinoleic acid by human peripheral blood eosinophils. Prostaglandins 1996, 52, 117–124. [Google Scholar] [CrossRef]
- Soberman, R.J.; Harper, T.W.; Betteridge, D.; Lewis, R.A.; Austen, K.F. Characterization and separation of the arachidonic acid 5-lipoxygenase and linoleic acid omega-6 lipoxygenase (arachidonic acid 15-lipoxygenase) of human polymorphonuclear leukocytes. J. Biol. Chem. 1985, 260, 4508–4515. [Google Scholar] [CrossRef]
- Kavadia, M.R.; Yadav, M.G.; Odaneth, A.A.; Lali, A.M. Production of glyceryl monostearate by immobilized candida antarctica B lipase in organic media. J. Appl. Biotechnol. Bioeng. 2017, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- van der Stelt, M.; Nieuwenhuizen, W.F.; Veldink, G.A.; Vliegenthart, J.F. Dioxygenation of N-linoleoyl amides by soybean lipoxygenase-1. FEBS Lett. 1997, 411, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, C.; Archambault, A.S.; Dumais, E.; Martin, C.; Blanchet, M.R.; Bissonnette, E.; Ohashi, N.; Yamamoto, K.; Itoh, T.; Laviolette, M.; et al. Endocannabinoid hydrolysis inhibition unmasks that unsaturated fatty acids induce a robust biosynthesis of 2-arachidonoyl-glycerol and its congeners in human myeloid leukocytes. FASEB J. 2020, 34, 4253–4265. [Google Scholar] [CrossRef] [Green Version]
- Archambault, A.S.; Poirier, S.; Lefebvre, J.S.; Robichaud, P.P.; Larose, M.C.; Turcotte, C.; Martin, C.; Provost, V.; Boudreau, L.H.; McDonald, P.P.; et al. 20-Hydroxy- and 20-carboxy-leukotriene (LT) B4 downregulate LTB4 -mediated responses of human neutrophils and eosinophils. J. Leukoc. Biol. 2019, 105, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Flamand, N.; Boudreault, S.; Picard, S.; Austin, M.; Surette, M.E.; Plante, H.; Krump, E.; Vallee, M.J.; Gilbert, C.; Naccache, P.; et al. Adenosine, a potent natural suppressor of arachidonic acid release and leukotriene biosynthesis in human neutrophils. Am. J. Respir. Crit. Care Med. 2000, 161, S88–S94. [Google Scholar] [CrossRef]
- Schmidt, M.W.; Baldridge, K.K.; Boatz, J.A.; Elbert, S.T.; Gordon, M.S.; Jensen, J.H.; Koseki, S.; Matsunaga, N.; Nguyen, K.A.; Su, S.; et al. General atomic and molecular electronic structure system. J. Comput. Chem. 1993, 14, 16. [Google Scholar]
- Fox, T.; Kollman, P.K. Application of the RESP Methodology in the Parametrization of Organic Solvents. J. Phys. Chem. 1998, 102, 9. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannotti, F.A.; De Maio, F.; Panza, E.; Appendino, G.; Taglialatela-Scafati, O.; De Petrocellis, L.; Amodeo, P.; Vitale, R.M. Identification and characterization of cannabimovone, a cannabinoid from cannabis sativa, as a novel ppargamma agonist via a combined computational and functional study. Molecules 2020, 25, 1119. [Google Scholar] [CrossRef] [Green Version]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.Y.; Brozellm, S.R.; Cerutti, D.S.; Cheatham, T.E.; Cisneros, G.A.; Cruzeiro, V.W.D.; Darden, T.A.; et al. AMBER 2016; University of California: San Francisco, CA, USA, 2016. [Google Scholar]
- Zoerner, A.A.; Gutzki, F.M.; Batkai, S.; May, M.; Rakers, C.; Engeli, S.; Jordan, J.; Tsikas, D. Quantification of endocannabinoids in biological systems by chromatography and mass spectrometry: A comprehensive review from an analytical and biological perspective. Biochim. Biophys. Acta 2011, 1811, 706–723. [Google Scholar] [CrossRef]
- Turcotte, C.; Dumais, E.; Archambault, A.S.; Martin, C.; Blanchet, M.R.; Bissonnette, E.; Boulet, L.P.; Laviolette, M.; Di Marzo, V.; Flamand, N. Human leukocytes differentially express endocannabinoid-glycerol lipases and hydrolyze 2-arachidonoyl-glycerol and its metabolites from the 15-lipoxygenase and cyclooxygenase pathways. J. Leukoc. Biol. 2019, 106, 1337–1347. [Google Scholar] [CrossRef]
- Deutsch, D.G.; Omeir, R.; Arreaza, G.; Salehani, D.; Prestwich, G.D.; Huang, Z.; Howlett, A. Methyl arachidonyl fluorophosphonate: A potent irreversible inhibitor of anandamide amidase. Biochem. Pharmacol. 1997, 53, 255–260. [Google Scholar] [CrossRef]
- Han, H.; Liang, X.; Ekberg, M.; Kritikou, J.S.; Brunnstrom, A.; Pelcman, B.; Matl, M.; Miao, X.; Andersson, M.; Yuan, X.; et al. Human 15-lipoxygenase-1 is a regulator of dendritic-cell spreading and podosome formation. FASEB J. 2017, 31, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Whitman, S.; Gezginci, M.; Timmermann, B.N.; Holman, T.R. Structure-activity relationship studies of nordihydroguaiaretic acid inhibitors toward soybean, 12-human, and 15-human lipoxygenase. J. Med. Chem 2002, 45, 2659–2661. [Google Scholar] [CrossRef]
- Tan, C.M.; Chen, G.S.; Chen, C.S.; Chang, P.T.; Chern, J.W. Design, synthesis and biological evaluation of benzo[1.3.2]dithiazolium ylide 1,1-dioxide derivatives as potential dual cyclooxygenase-2/5-lipoxygenase inhibitors. Bioorg. Med. Chem. 2011, 19, 6316–6328. [Google Scholar] [CrossRef]
- Suriano, F.; Manca, C.; Flamand, N.; Depommier, C.; Van Hul, M.; Delzenne, N.M.; Silvestri, C.; Cani, P.D.; Di Marzo, V. Exploring the endocannabinoidome in genetically obese (ob/ob) and diabetic (db/db) mice: links with inflammation and gut microbiota. Biochim. Biophys. Acta Mol. Cell Biol Lipids 2021. under review. [Google Scholar]
- Urquhart, P.; Nicolaou, A.; Woodward, D.F. Endocannabinoids and their oxygenation by cyclo-oxygenases, lipoxygenases and other oxygenases. Biochim. Biophys. Acta 2015, 1851, 366–376. [Google Scholar] [CrossRef]
- Ivanov, I.; Kakularam, K.R.; Shmendel, E.V.; Rothe, M.; Aparoy, P.; Heydeck, D.; Kuhn, H. Oxygenation of endocannabinoids by mammalian lipoxygenase isoforms. Biochim. Biophys. Acta Mol. Cell Biol Lipids 2021, 1866, 158918. [Google Scholar] [CrossRef] [PubMed]
- Upston, J.M.; Neuzil, J.; Witting, P.K.; Alleva, R.; Stocker, R. Oxidation of free fatty acids in low density lipoprotein by 15-lipoxygenase stimulates nonenzymic, alpha-tocopherol-mediated peroxidation of cholesteryl esters. J. Biol. Chem. 1997, 272, 30067–30074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammerer, I.; Ringseis, R.; Biemann, R.; Wen, G.; Eder, K. 13-hydroxy linoleic acid increases expression of the cholesterol transporters ABCA1, ABCG1 and SR-BI and stimulates apoA-I-dependent cholesterol efflux in RAW264.7 macrophages. Lipids Health Dis. 2011, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.W.; Balzar, S.; Westcott, J.Y.; Trudeau, J.B.; Sun, Y.; Conrad, D.J.; Wenzel, S.E. Expression and activation of 15-lipoxygenase pathway in severe asthma: Relationship to eosinophilic phenotype and collagen deposition. Clin. Exp. Allergy 2002, 32, 1558–1565. [Google Scholar] [CrossRef]
- Singh, N.K.; Rao, G.N. Emerging role of 12/15-Lipoxygenase (ALOX15) in human pathologies. Prog. Lipid Res. 2019, 73, 28–45. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid | Internal Standard Used | Q1 → Q3 | Retention Time (Min) | LLOQ (Fmol) |
|---|---|---|---|---|
| 13-HODE-d4 | - | 299.10 → 198.15 | 9.89 | - |
| 13-HODE-G-d5 | - | 358.00 → 261.30 | 8.76 | - |
| 13-HODE-EA-d4 | - | 326.30 → 66.20 | 7.72 | - |
| 13-HODE | 13-HODE-d4 | 295.5 → 277.30 | 9.94 | 25 |
| 13-HODE-G | 13-HODE-G-d5 | 353.20 → 261.2 | 8.80 | 25 |
| 13-HODE-EA | 13-HODE-EA-d4 | 322.00 → 62.2 | 7.76 | 25 |
| CARBON | 13-HODE-G (2) | |||
|---|---|---|---|---|
| N° | δ C (ppm) | δ H (ppm) | Features | J (Hz) |
| 1 | 174.3 | NA | NA | NA |
| 2 | 34.1 | 2.35 | t | 7.5 |
| 3 | 24.8 | 1.67–1.47 | 2H, br m | |
| 4 | 22.6 | 1.32 | m | |
| 5 | 28.9 | 1.32 | m | |
| 6 | 28.9 | 1.32 | m | |
| 7 | 28.9 | 1.32 | m | |
| 8 | 27.6 | 2.18 | m | |
| 9 | 132.8 | 5.44 | m | |
| 10 | 127.8 | 5.98 | t | 10.9 |
| 11 | 125.7 | 6.49 | dd | 15.2, 11.0 |
| 12 | 135.9 | 5.67 | dd | 15.2, 6.8 |
| 13 | 72.9 | 4.16 | 1H, overlapped m | |
| 14 | 37.3 | 1.67–1.47 | 2H, br m | |
| 15 | 29.7 | 1.32 | m | |
| 16 | 29.3 | 1.32 | m | |
| 17 | 25.1 | 1.32 | m | |
| 18 | 14.1 | 0.89 | t | 7 |
| 1’ | 70.3 | 3.93 | 1H, m | |
| 2’ | 65.2 | 4.16 | 2H, overlapped m | |
| 3’ | 63.3 | 3.65 | 2H, m | |
| Duodenum | Jejunum | Ileum | Caecum |
|---|---|---|---|
| 9746 ± 4360 | 4810 ± 2692 | 1165 ± 185 | 997 ± 317 |
| Compound | CB1 Receptor | CB2 Receptor | ||||
|---|---|---|---|---|---|---|
| IC50 (µM) | Ki (µM) | Max Tested (% Displacement) | IC50 (µM) | Ki (µM) | Max Tested (% Displacement) | |
| 13-HODE | >10 | >10 | 10 µM (7.56 ± 2.3) | >10 | >10 | 10 µM (38.34 ± 5.4) |
| 13-HODE-G | >10 | >10 | 10 µM (18.90 ± 6.1) | >10 | >10 | 10 µM (33.58 ± 21.3) |
| 1-AG | 1.32 ± 0.46 | 0.15 ± 0.05 | 10 µM (90.4 ± 1.3) | 1.02 ± 0.04 | 0.16 ± 0.01 | 10 µM (75.52 ± 2.4) |
| 1-LG | 2.81 ± 0.85 | 0.31 ± 0.15 | 10µM (95.8 ± 16.4) | >10 | >10 | 10 µM (28.45 ± 9.4) |
| Compound | Efficacy (% Max Response) | Potency (EC50 in µM) | Desensibilization 1 (IC50 in µM) |
|---|---|---|---|
| 1-AG | 58.9 ± 1.2 | 0.39 ± 0.03 | 1.1 ± 0.1 |
| 1-LG | 46.5 ± 0.9 | 0.23 ± 0.02 | 2.7 ± 0.3 |
| 13-HODE | < 10 | - | > 50 |
| 13-HODE-G | < 10 | - | > 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Archambault, A.-S.; Tinto, F.; Dumais, É.; Rakotoarivelo, V.; Kostrzewa, M.; Plante, P.-L.; Martin, C.; Simard, M.; Silvestri, C.; Pouliot, R.; et al. Biosynthesis of the Novel Endogenous 15-Lipoxygenase Metabolites N-13-Hydroxy-octodecadienoyl-ethanolamine and 13-Hydroxy-octodecadienoyl-glycerol by Human Neutrophils and Eosinophils. Cells 2021, 10, 2322. https://doi.org/10.3390/cells10092322
Archambault A-S, Tinto F, Dumais É, Rakotoarivelo V, Kostrzewa M, Plante P-L, Martin C, Simard M, Silvestri C, Pouliot R, et al. Biosynthesis of the Novel Endogenous 15-Lipoxygenase Metabolites N-13-Hydroxy-octodecadienoyl-ethanolamine and 13-Hydroxy-octodecadienoyl-glycerol by Human Neutrophils and Eosinophils. Cells. 2021; 10(9):2322. https://doi.org/10.3390/cells10092322
Chicago/Turabian StyleArchambault, Anne-Sophie, Francesco Tinto, Élizabeth Dumais, Volatiana Rakotoarivelo, Magdalena Kostrzewa, Pier-Luc Plante, Cyril Martin, Mélissa Simard, Cristoforo Silvestri, Roxane Pouliot, and et al. 2021. "Biosynthesis of the Novel Endogenous 15-Lipoxygenase Metabolites N-13-Hydroxy-octodecadienoyl-ethanolamine and 13-Hydroxy-octodecadienoyl-glycerol by Human Neutrophils and Eosinophils" Cells 10, no. 9: 2322. https://doi.org/10.3390/cells10092322