


Parsimonious Effect of Pentoxifylline on Angiogenesis: A Novel Pentoxifylline-Biased Adenosine G Protein-Coupled Receptor Signaling Platform
 , , and
, , and
Abstract
:1. Introduction
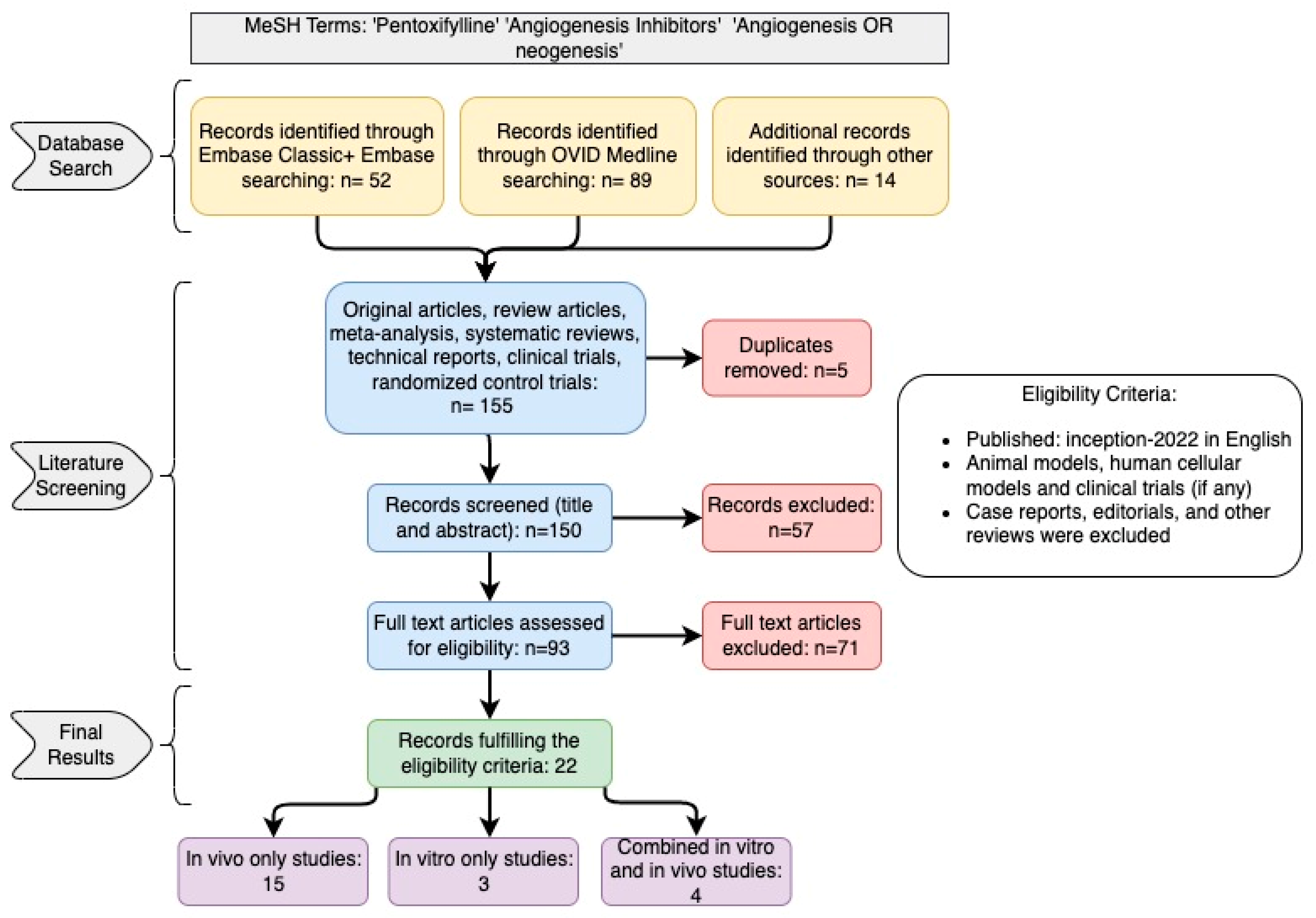
2. Study Design and Search Strategy
3. Results
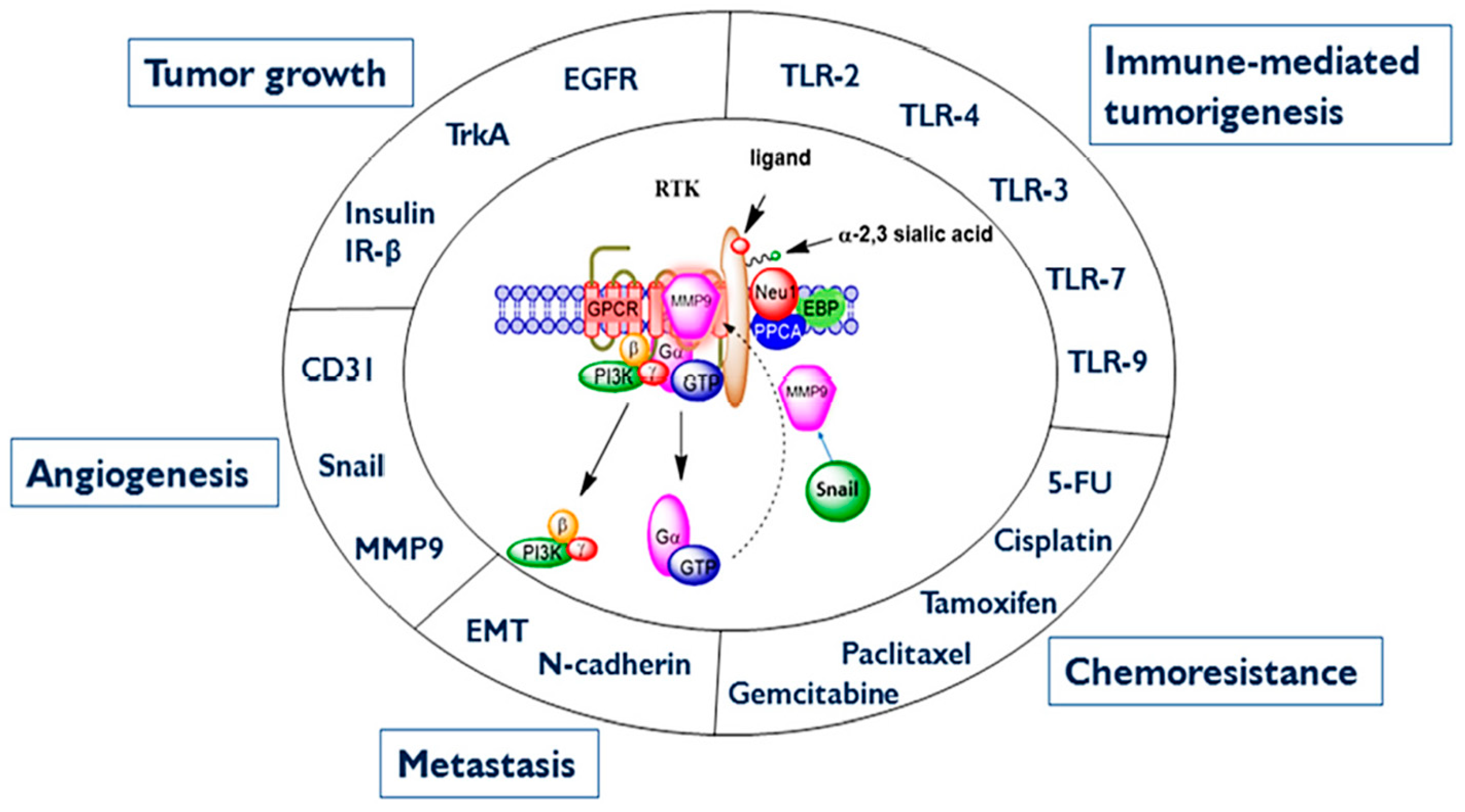
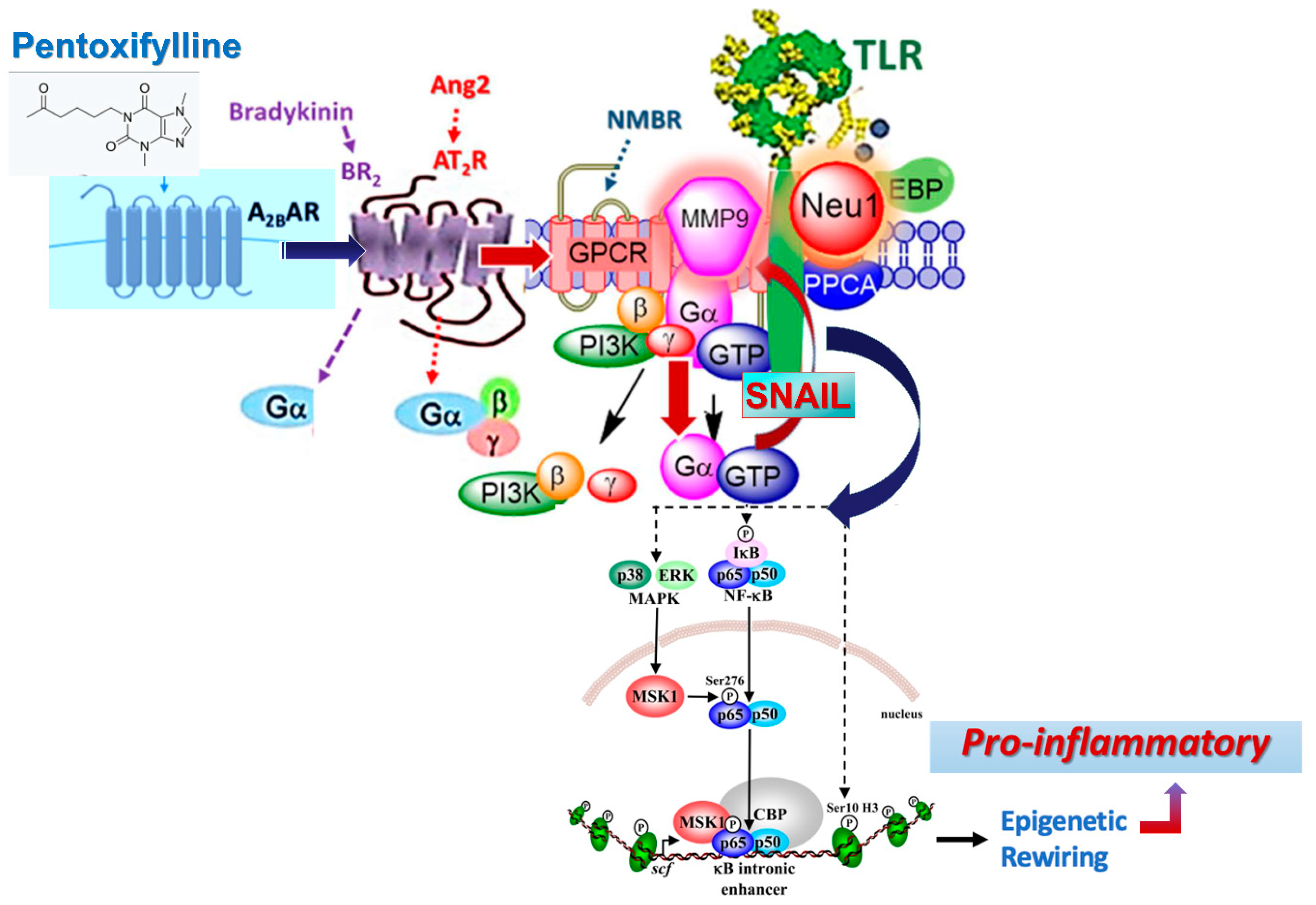
4. Discussion

5. Limitations of the Study
6. Conclusions
7. Future Research Directions and Limitations
- Exploring the structure–activity relationships of PTX that target the adenosine GPCR site or behave as neutral A2B adenosine receptor (A2BAR) antagonists.
- Assessing the therapeutic potential of pentoxifylline-biased A2BAR receptor allosteric modulators and neutral antagonists.
- Validating and characterizing other targets, such as RTK and TLR receptors, for particular PTX effects and developing compounds in selectively activating or blocking targets with potency for metabolic health and related diseases.
- Validating and characterizing A2B adenosine receptors as homodimers, heterodimers, or oligomers with one or more classes of other receptors.
- Validating and characterizing the angiogenesis system in ameliorating the symptoms or the underlying pathology of certain disorders.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FGF | Fibroblast growth factor |
| IL | Interleukin |
| MMP | Matrix metalloproteinase |
| PDGF | Platelet-derived growth factor |
| SCF | Stem cell factor |
| STAT3 | Signal transducer and activator of transcription 3 |
| TNF | Tumor necrosis factor |
| VEGF | Vascular endothelial growth factor |
| PECAM | Platelet endothelial cell adhesion molecule |
| TGF | Tumor growth factor |
| HIF | Hypoxia-inducible factor |
| DMXAA | Dimethylxanthenone-4-acetic acid |
References
- Eelen, G.; Treps, L.; Li, X.; Carmeliet, P. Basic and Therapeutic Aspects of Angiogenesis Updated. Circ. Res. 2020, 127, 310–329. [Google Scholar] [CrossRef]
- Marech, I.; Leporini, C.; Ammendola, M.; Porcelli, M.; Gadaleta, C.D.; Russo, E.; De Sarro, G.; Ranieri, G. Classical and non-classical proangiogenic factors as a target of antiangiogenic therapy in tumor microenvironment. Cancer Lett. 2016, 380, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Fundamental concepts of the angiogenic process. Curr. Mol. Med. 2003, 3, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.; Satchi-Fainaro, R. Design and development of polymer conjugates as anti-angiogenic agents. Adv. Drug Deliv. Rev. 2009, 61, 1159–1176. [Google Scholar] [CrossRef]
- Kerbel, R.S.; Kamen, B.A. The anti-angiogenic basis of metronomic chemotherapy. Nat. Rev. Cancer 2004, 4, 423–436. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, Y.J.; Mengi, S.A.; Arneja, A.S.; Dhalla, N.S. Therapeutic potentials of pentoxifylline for treatment of cardiovascular diseases. Exp. Clin. Cardiol. 2004, 9, 103–111. [Google Scholar]
- Salhiyyah, K.; Forster, R.; Senanayake, E.; Abdel-Hadi, M.; Booth, A.; Michaels, J.A. Pentoxifylline for intermittent claudication. Cochrane Database Syst. Rev. 2015, 9, Cd005262. [Google Scholar] [CrossRef]
- Fernandes, J.L.; de Oliveira, R.T.D.; Mamoni, R.L.; Coelho, O.R.; Nicolau, J.C.; Blotta, M.; Serrano, C.V., Jr. Pentoxifylline reduces pro-inflammatory and increases anti-inflammatory activity in patients with coronary artery disease—A randomized placebo-controlled study. Atherosclerosis 2008, 196, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Herskovits, E.; Famulari, A.; Tamaroff, L.; Gonzalez, A.M.; Vázquez, A.; Dominguez, R.; Fraiman, H.; Vila, J. Preventive treatment of cerebral transient ischemia: Comparative randomized trial of pentoxifylline versus conventional antiaggregants. Eur. Neurol. 1985, 24, 73–81. [Google Scholar] [CrossRef]
- Champion, S.; Lapidus, N.; Cherié, G.; Spagnoli, V.; Oliary, J.; Solal, A.C. Pentoxifylline in heart failure: A meta-analysis of clinical trials. Cardiovasc. Ther. 2014, 32, 159–162. [Google Scholar] [CrossRef]
- Berman, B.; Duncan, M.R. Pentoxifylline inhibits normal human dermal fibroblast in vitro proliferation, collagen, glycosaminoglycan, and fibronectin production, and increases collagenase activity. J. Investig. Dermatol. 1989, 92, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, U.; Ukena, D.; Lohse, M.J. Xanthine derivatives as antagonists at A1 and A2 adenosine receptors. Naunyn Schmiedebergs Arch. Pharmacol. 1985, 330, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Kreth, S.; Ledderose, C.; Luchting, B.; Weis, F.; Thiel, M. Immunomodulatory properties of pentoxifylline are mediated via adenosine-dependent pathways. Shock 2010, 34, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Konrad, F.M.; Neudeck, G.; Vollmer, I.; Ngamsri, K.C.; Thiel, M.; Reutershan, J. Protective effects of pentoxifylline in pulmonary inflammation are adenosine receptor A2A dependent. Faseb J. 2013, 27, 3524–3535. [Google Scholar] [CrossRef]
- Li, H.; Tan, G.; Tong, L.; Han, P.; Zhang, F.; Liu, B.; Sun, X. Pentoxifylline inhibits pulmonary inflammation induced by infrarenal aorticcross-clamping dependent of adenosine receptor A2A. Am. J. Transl. Res. 2016, 8, 2210–2221. [Google Scholar]
- Pyne, N.J.; Waters, C.M.; Long, J.S.; Moughal, N.A.; Tigyi, G.; Pyne, S. Receptor tyrosine kinase-G-protein coupled receptor complex signaling in mammalian cells. Adv. Enzym. Regul. 2007, 47, 271–280. [Google Scholar] [CrossRef]
- Pyne, N.J.; Pyne, S. Receptor tyrosine kinase-G-protein-coupled receptor signalling platforms: Out of the shadow? Trends Pharmacol. Sci. 2011, 32, 443–450. [Google Scholar] [CrossRef]
- Pyne, N.J.; Waters, C.; Moughal, N.A.; Sambi, B.S.; Pyne, S. Receptor tyrosine kinase-GPCR signal complexes. Biochem. Soc. Trans. 2003, 31, 1220–1225. [Google Scholar] [CrossRef]
- Abdulkhalek, S.H.M.; Szewczuk, M.R. A novel G-protein-coupled receptor-signaling platform and its targeted translation in human disease. Res. Rep. Biochem. 2013, 3, 17–30. [Google Scholar]
- Haxho, F.A.F.; Neufeld, R.J.; Szewczuk, M.R. Novel insulin receptor signaling platform. Int. J. Diabetes Clin. Res. 2014, 1, 5. [Google Scholar] [CrossRef]
- Onfroy, L.; Galandrin, S.; Pontier, S.M.; Seguelas, M.H.; N′Guyen, D.; Sénard, J.M.; Galés, C. G protein stoichiometry dictates biased agonism through distinct receptor-G protein partitioning. Sci. Rep. 2017, 7, 7885. [Google Scholar] [CrossRef] [PubMed]
- Khoury, E.; Clément, S.; Laporte, S.A. Allosteric and biased g protein-coupled receptor signaling regulation: Potentials for new therapeutics. Front. Endocrinol. 2014, 5, 68. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, L.M.; Maudsley, S.; Bohn, L.M. Fulfilling the Promise of “Biased” G Protein-Coupled Receptor Agonism. Mol. Pharmacol. 2015, 88, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.R.; May, L.T.; Parton, R.G.; Sexton, P.M.; Christopoulos, A. A kinetic view of GPCR allostery and biased agonism. Nat. Chem. Biol. 2017, 13, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, S.J.; Changeux, J.P. Biased Allostery. Biophys. J. 2016, 111, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, J.L.; Ambrus, C.M.; Toumbis, C.A.; Forgach, P.; Karakousis, C.P.; Niswander, P.; Lane, W. Studies on tumor induced angiogenesis. J. Med. 1991, 22, 355–369. [Google Scholar]
- Ambrus, J.L.; Ambrus, C.M.; Forgach, P.; Stadler, S.; Halpern, J.; Sayyid, S.; Niswander, P.; Toumbis, C. Studies on tumor induced angiogenesis. Exp. Suppl. 1992, 61, 436–444. [Google Scholar] [CrossRef]
- Ambrus, J.L.; Toumbis, C.A.; Karakousis, C.P.; Kulaylat, M.; Akhter, S.; Plavsic, L. Study of antiangiogenic agents with possible therapeutic applications in neoplastic disorders and macular degeneration. J. Med. 2000, 31, 278–282. [Google Scholar]
- Ching, L.M.; Browne, W.L.; Tchernegovski, R.; Gregory, T.; Baguley, B.C.; Palmer, B.D. Interaction of thalidomide, phthalimide analogues of thalidomide and pentoxifylline with the anti-tumour agent 5,6-dimethylxanthenone-4-acetic acid: Concomitant reduction of serum tumour necrosis factor-alpha and enhancement of anti-tumour activity. Br. J. Cancer 1998, 78, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Luo, B.; Tang, L.; Wang, Y.; Stockard, C.R.; Kadish, I.; Van Groen, T.; Grizzle, W.E.; Ponnazhagan, S.; Fallon, M.B. Pulmonary angiogenesis in a rat model of hepatopulmonary syndrome. Gastroenterology 2009, 136, 1070–1080. [Google Scholar] [CrossRef]
- Mendes, J.B.; Campos, P.P.; Rocha, M.A.; Andrade, S.P. Cilostazol and pentoxifylline decrease angiogenesis, inflammation, and fibrosis in sponge-induced intraperitoneal adhesion in mice. Life Sci. 2009, 84, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Vlahos, N.F.; Gregoriou, O.; Deliveliotou, A.; Perrea, D.; Vlachos, A.; Zhao, Y.; Lai, J.; Creatsas, G. Effect of pentoxifylline on vascular endothelial growth factor C and flk-1 expression on endometrial implants in the rat endometriosis model. Fertil. Steril. 2010, 93, 1316–1323. [Google Scholar] [CrossRef]
- Boztosun, A.; Piçnak, A.; Kosar, M.I.; Gulturk, S.; Cetin, A. Effects of methylene blue, pentoxyphylline and enoxaparin on postoperative adhesion formation and markers of angiogenesis in a rat uterine horn model. Clin. Exp. Obstet. Gynecol. 2012, 39, 89–95. [Google Scholar]
- Pratibha, D.Y.N.; Rajiv, G. Pentoxifylline: A Potent Inhibitor of Angiogenesis via Blocking STAT3 Signaling in B16F10 Melanoma. Int. J. Tumor. Ther. 2013, 2, 1–9. [Google Scholar] [CrossRef]
- Çakmak, G.; Şahin, M.; Özdemİr, B.H.; Karadenİz, E. Effect of pentoxifylline on healing of segmental bone defects and angiogenesis. Acta Orthop. Traumatol. Turc. 2015, 49, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Nathan, J.R.; Lakshmanan, G.; Michael, F.M.; Seppan, P.; Ragunathan, M. Expression of adenosine receptors and vegf during angiogenesis and its inhibition by pentoxifylline-A study using zebrafish model. Biomed. Pharmacother. 2016, 84, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Bałan, B.J.; Demkow, U.; Skopiński, P.; Bychawska, M.; Skopińska-Różewska, E.; Lewicki, S.; Zdanowski, R. The effect of pentoxifylline on L-1 sarcoma tumor growth and angiogenesis in Balb/c mice. Cent. Eur. J. Immunol. 2017, 42, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Lee, M.G.; Lee, C.C.; Su, P.I.; Chi, C.Y.; Liu, C.H.; Wu, M.C.; Yen, Z.S.; Chen, S.C. Pentoxifylline decreases post-operative intra-abdominal adhesion formation in an animal model. PeerJ 2018, 6, e5434. [Google Scholar] [CrossRef]
- Pedretti, S.; Rena, C.L.; Orellano, L.A.A.; Lazari, M.G.; Campos, P.P.; Nunes, T.A. Benefits of pentoxifylline for skin flap tissue repair in rats. Acta Cir. Bras. 2020, 35, e301105. [Google Scholar] [CrossRef]
- Seo, M.H.; Myoung, H.; Lee, J.H.; Yang, H.C.; Woo, K.M.; Lee, S.K.; Kim, S.M. Effects of pentoxifylline and tocopherol on an osteoradionecrosis animal model. J. Craniomaxillofac. Surg. 2020, 48, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Niderla-Bielińska, J.; Bartkowiak, K.; Ciszek, B.; Czajkowski, E.; Jankowska-Steifer, E.; Krejner, A.; Ratajska, A. Pentoxifylline inhibits angiogenesis via decreasing Dll4 and Notch1 expression in mouse proepicardial explant cultures. Eur. J. Pharmacol. 2018, 827, 80–87. [Google Scholar] [CrossRef]
- Arsenyan, P.V.J.; Domracheva, I.; Kanepe-Lapsa, I. 8-Ethynylxanthines as promising antiproliferative agents, angiogenesis inhibitors, and calcium channel activity modulators. Chem. Heterocycl. Compd. 2020, 56, 776–785. [Google Scholar] [CrossRef]
- Seo, M.H.; Eo, M.Y.; Nguyen, T.T.H.; Yang, H.J.; Kim, S.M. Immunomodulatory Effects of Pentoxifylline: Profiling Data Based on RAW 264.7 Cellular Signaling. Appl. Sci. 2021, 11, 8273. [Google Scholar] [CrossRef]
- Joseph, I.B.; Isaacs, J.T. Macrophage role in the anti-prostate cancer response to one class of antiangiogenic agents. J. Natl. Cancer Inst. 1998, 90, 1648–1653. [Google Scholar] [CrossRef] [PubMed]
- Gude, R.P.; Binda, M.M.; Boquete, A.L.; Bonfil, R.D. Inhibition of endothelial cell proliferation and tumor-induced angiogenesis by pentoxifylline. J. Cancer Res. Clin. Oncol. 2001, 127, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.Z.; Gude, R.P. Pentoxifylline inhibits melanoma tumor growth and angiogenesis by targeting STAT3 signaling pathway. Biomed. Pharmacother. 2013, 67, 399–405. [Google Scholar] [CrossRef]
- Nidhyanandan, S.; Boreddy, T.S.; Chandrasekhar, K.B.; Reddy, N.D.; Kulkarni, N.M.; Narayanan, S. Phosphodiesterase inhibitor, pentoxifylline enhances anticancer activity of histone deacetylase inhibitor, MS-275 in human breast cancer in vitro and in vivo. Eur. J. Pharmacol. 2015, 764, 508–519. [Google Scholar] [CrossRef]
- Smith, R.V.; Waller, E.S.; Doluisio, J.T.; Bauza, M.T.; Puri, S.K.; Ho, I.; Lassman, H.B. Pharmacokinetics of orally administered pentoxifylline in humans. J. Pharm. Sci. 1986, 75, 47–52. [Google Scholar] [CrossRef]
- Ward, A.; Clissold, S.P. Pentoxifylline. Drugs 1987, 34, 50–97. [Google Scholar] [CrossRef]
- D'Hellencourt, C.L.; Diaw, L.; Cornillet, P.; Guenounou, M. Differential regulation of TNF alpha, IL-1 beta, IL-6, IL-8, TNF beta, and IL-10 by pentoxifylline. Int. J. Immunopharmacol. 1996, 18, 739–748. [Google Scholar] [CrossRef]
- Strieter, R.M.; Remick, D.G.; Ward, P.A.; Spengler, R.N.; Lynch, J.P., 3rd; Larrick, J.; Kunkel, S.L. Cellular and molecular regulation of tumor necrosis factor-alpha production by pentoxifylline. Biochem. Biophys. Res. Commun. 1988, 155, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Neuner, P.; Klosner, G.; Schauer, E.; Pourmojib, M.; Macheiner, W.; Grünwald, C.; Knobler, R.; Schwarz, A.; Luger, T.; Schwarz, T. Pentoxifylline in vivo down-regulates the release of IL-1 beta, IL-6, IL-8 and tumour necrosis factor-alpha by human peripheral blood mononuclear cells. Immunology 1994, 83, 262. [Google Scholar] [PubMed]
- Mendes, J.B.; Rocha, M.A.; Araújo, F.A.; Moura, S.A.; Ferreira, M.A.; Andrade, S.P. Differential effects of rolipram on chronic subcutaneous inflammatory angiogenesis and on peritoneal adhesion in mice. Microvasc. Res. 2009, 78, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Samlaska, C.P.; Winfield, E.A. Pentoxifylline. J. Am. Acad. Dermatol. 1994, 30, 603–621. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef]
- Panneerselvan, N.; Ragunathan, M. Targeting expression of adenosine receptors during hypoxia induced angiogenesis - A study using zebrafish model. Biomed. Pharmacother. 2018, 99, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Bessa-Gonçalves, M.; Bragança, B.; Martins-Dias, E.; Correia-de-Sa, P.; Fontes-Sousa, A.P. Is the adenosine A2B ‘biased’receptor a valuable target for the treatment of pulmonary arterial hypertension? Drug Discov. Today 2018, 23, 1285–1292. [Google Scholar] [CrossRef]
- Effendi, W.I.; Nagano, T.; Kobayashi, K.; Nishimura, Y. Focusing on adenosine receptors as a potential targeted therapy in human diseases. Cells 2020, 9, 785. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Frenguelli, B.G.; Hills, R.; IJzerman, A.P.; Jacobson, K.A.; Klotz, K.-N.; Linden, J.; Müller, C.E.; Schwabe, U.; Stiles, G.L. Adenosine receptors in GtoPdb. IUPHAR/BPS Guide Pharmacol. 2021, 2021. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Synowitz, M.; Powell, J.D.; Klotz, K.N.; Gessi, S.; Borea, P.A. Adenosine receptors and cancer. Handb. Exp. Pharmacol. 2009, 193, 399–441. [Google Scholar] [CrossRef]
- D′Antongiovanni, V.; Fornai, M.; Pellegrini, C.; Benvenuti, L.; Blandizzi, C.; Antonioli, L. The Adenosine System at the Crossroads of Intestinal Inflammation and Neoplasia. Int. J. Mol. Sci. 2020, 21, 5089. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, A.M.; Abdulkhalek, S.; Cheng, T.S.; Alghamdi, F.; Jayanth, P.; O’Shea, L.K.; Geen, O.; Arvizu, L.A.; Szewczuk, M.R. A novel epidermal growth factor receptor-signaling platform and its targeted translation in pancreatic cancer. Cell. Signal. 2013, 25, 2587–2603. [Google Scholar] [CrossRef] [PubMed]
- Haxho, F.; Neufeld, R.J.; Szewczuk, M.R. Neuraminidase-1: A novel therapeutic target in multistage tumorigenesis. Oncotarget 2016, 7, 40860. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhalek, S.; Geen, O.D.; Brodhagen, L.; Haxho, F.; Alghamdi, F.; Allison, S.; Simmons, D.J.; O’Shea, L.K.; Neufeld, R.J.; Szewczuk, M.R. Transcriptional factor snail controls tumor neovascularization, growth and metastasis in mouse model of human ovarian carcinoma. Clin. Transl. Med. 2014, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhalek, S.; Guo, M.; Amith, S.R.; Jayanth, P.; Szewczuk, M.R. G-protein coupled receptor agonists mediate Neu1 sialidase and matrix metalloproteinase-9 crosstalk to induce transactivation of TOLL-like receptors and cellular signaling. Cell. Signal. 2012, 24, 2035–2042. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Verma, I.M. Nuclear factor (NF)-κB proteins: Therapeutic targets. Ann. Rheum. Dis. 2004, 63, ii57. [Google Scholar] [CrossRef]
- Berera, S.; Ioannou, S.C.; Morillo, D.; Mantero, A.M.A.; Pignac-Kobinger, J.; Colina, N.; Santander, A.M.; Fernandez, I.; Quintero, M.A.; Rodriguez, J.; et al. Combining Pentoxifylline With Vedolizumab for Crohn’s Disease: Results of a Randomised, Placebo-controlled Pilot Study. J. Crohns Colitis 2022, 16, 1687–1695. [Google Scholar] [CrossRef]
- Vege, S.S.; Atwal, T.; Bi, Y.; Chari, S.T.; Clemens, M.A.; Enders, F.T. Pentoxifylline Treatment in Severe Acute Pancreatitis: A Pilot, Double-Blind, Placebo-Controlled, Randomized Trial. Gastroenterology 2015, 149, 318–320.e313. [Google Scholar] [CrossRef]
- Saki, N.; Sheikhi Ghayur, E.; Heiran, A.; Gholami, M.; Alipour, S. Topical pentoxifylline can be an effective and safe adjunctive therapy to NBUVB therapy in treating vitiligo: A split-side clinical trial. Exp. Dermatol. 2022, 31, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, H.J.N.; Matthews, J.; Adams, C.; Gothard, L.; Lucy, C.; Tovey, H.; Boyle, S.; Anbalagan, S.; Musallam, A.; Yarnold, J.; et al. Randomised single centre double-blind placebo controlled phase II trial of Tocovid SupraBio in combination with pentoxifylline in patients suffering long-term gastrointestinal adverse effects of radiotherapy for pelvic cancer: The PPALM study. Radiother. Oncol. 2022, 168, 130–137. [Google Scholar] [CrossRef]
- Ozturk, B.; Egehan, I.; Atavci, S.; Kitapci, M. Pentoxifylline in prevention of radiation-induced lung toxicity in patients with breast and lung cancer: A double-blind randomized trial. Int. J. Radiat. Oncol. Biol. Phys. 2004, 58, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Meirovitz, A.; Baider, L.; Peretz, T.; Stephanos, S.; Barak, V. PTX Treatment of Colon Cancer: Mode of Action Based on Tumor Marker and Cytokine Kinetics. Anticancer Res. 2022, 42, 5487–5496. [Google Scholar] [CrossRef]
- Misirlioglu, C.H.; Erkal, H.; Elgin, Y.; Ugur, I.; Altundag, K. Effect of concomitant use of pentoxifylline and alpha-tocopherol with radiotherapy on the clinical outcome of patients with stage IIIB non-small cell lung cancer: A randomized prospective clinical trial. Med. Oncol. 2006, 23, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.J.; Cripps, M.C.; Fingert, H.; Lamothe, A.; Laframboise, G.; Gerin-Lajoie, J. Cisplatin, cytosine arabinoside, and pentoxifylline in the treatment of squamous cell carcinoma of the head and neck. Am. J. Clin. Oncol. 1993, 16, 123–126. [Google Scholar] [CrossRef]
- Ott, E.; Lechner, H.; Fazekas, F. Hemorheological effects of pentoxifylline on disturbed flow behavior of blood in patients with cerebrovascular insufficiency. Eur. Neurol. 1983, 22 (Suppl. 1), 105–107. [Google Scholar] [CrossRef]
- Oh, S.P.; Seki, T.; Goss, K.A.; Imamura, T.; Yi, Y.; Donahoe, P.K.; Li, L.; Miyazono, K.; ten Dijke, P.; Kim, S.; et al. Activin receptor-like kinase 1 modulates transforming growth factor-beta 1 signaling in the regulation of angiogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2626–2631. [Google Scholar] [CrossRef]
- Qorri, B.; Kalaydina, R.V.; Velickovic, A.; Kaplya, Y.; Decarlo, A.; Szewczuk, M.R. Agonist-Biased Signaling via Matrix Metalloproteinase-9 Promotes Extracellular Matrix Remodeling. Cells 2018, 7, 117. [Google Scholar] [CrossRef]
- Jakowiecki, J.; Abel, R.; Orzeł, U.; Pasznik, P.; Preissner, R.; Filipek, S. Allosteric Modulation of the CB1 Cannabinoid Receptor by Cannabidiol-A Molecular Modeling Study of the N-Terminal Domain and the Allosteric-Orthosteric Coupling. Molecules 2021, 26, 2456. [Google Scholar] [CrossRef]
- Reber, L.; Vermeulen, L.; Haegeman, G.; Frossard, N. Ser276 phosphorylation of NF-kB p65 by MSK1 controls SCF expression in inflammation. PLoS ONE 2009, 4, e4393. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Study [Reference] | Study Design (In Vivo) | Study Design (In Vitro) | Findings |
|---|---|---|---|
| Ambrus et al., 1991 [26] | Human malignant melanoma implants in rabbit cornea and non-human primates. | Pentoxifylline inhibited human tumor implant-induced angiogenesis. | |
| Ambrus et al., 1992 [27] | Human malignant melanoma implants in rabbit cornea and non-human primates. | Pentoxifylline inhibited human tumor implant-induced angiogenesis. | |
| Ching et al., 1998 [29] | Interaction of thalidomide, phthalimide analogs of thalidomide and pentoxifylline with anti-tumor agent DMXAA in mice models given Colon 38 tumors. | Pentoxifylline potentiates DMXAA inhibition of serum TNF production. | |
| Joseph & Isaacs, 1998 [44] | Transplanted Dunning R-3327 MAT-Lu rat prostate cancers. Pentoxifylline effects were assessed on tumor-associated macrophages and blood vessel densities. | On human and mouse macrophages. | Pentoxifylline inhibition of macrophage secretion tumor necrosis factor-α and granulocyte–macrophage colony-stimulating factor, and reduction of tumor blood vessel density and tumor growth. |
| Ambrus et al., 2000 [28] | Human malignant melanoma implants in Macaca arctoides monkey cornea. | Pentoxifylline inhibited human tumor implant-induced angiogenesis. | |
| Gude et al., 2001 [45] | C57BL/6J mice model injected with B16-F10 melanoma cells. IC50 7 mM after 24 h exposure. | Growth inhibition of 2 endothelial cell lines. | Significant inhibition of tumor-induced angiogenesis in C57B1/6 mice inoculated with pentoxifylline that paralleled decreased tumor volumes. In vitro pentoxifylline exhibited a dose-response inhibition of endothelial cell growth and downregulation of urokinase-type plasminogen activator expression. |
| Zhang et al., 2009 [30] | Hepatopulmonary syndrome induced in a rat model; pulmonary angiogenesis was assessed by quantifying factor VIII-positive micro vessels and levels of von Willebrand factor, vascular endothelial cadherin, angiogenic factors, and proliferating cell nuclear antigen. | Pentoxifylline-treated rats had a reduction in micro vessels and lung monocyte accumulation, downregulation of pulmonary angiogenic factors, and decreased symptoms of hepatopulmonary syndrome. The authors conclude that pentoxifylline decreases hepatopulmonary syndrome-associated angiogenesis, decreases the associated symptoms, and downregulates VEGF-A mediated pathways. | |
| Mendes et al., 2009 [31] | Murine model of sponge-induced peritoneal adhesion, treated with pentoxifylline and assessed by measuring hemoglobin content, VEGF, and morphometric analysis. | Following treatments of pentoxifylline, hemoglobin content, morphometric, morphometric analysis of vessel number, and levels of VEGF decreased. The results align with previous evidence that anti-VEGF activity is associated with angiogenesis inhibition. | |
| Vlahos et al., 2010 [32] | Surgical induction of endometriosis in rats. Morphological changes and VEGF-C and FLK-1 expression were assessed. | There was a significant reduction in the mean volume of endometriotic implants in the pentoxifylline treatment groups. There was a significant reduction in VEGF-C and FLK-1 expressions. The authors conclude that pentoxifylline may suppress angiogenesis by downregulating VEGF-C and FLK-1 expression. | |
| Boztosun et al., 2012 [33] | Surgical induction of adhesions in rats. Morphological changes and VEGF, bFGF, TGF-β, and PDGF expression were assessed. | Pentoxifylline did not show any effect on the expression of angiogenic factors. | |
| Pratibha et al., 2013 [34] | This study investigated the mechanisms for the antiangiogenic activity of pentoxifylline by injecting B16-F10 melanoma cells into C57BL/6 mice and assessing blood vessel density and molecular markers. IC50 39.2 +/− 1.3 mM after 2 h exposure. | The results of this study demonstrated that pentoxifylline: suppressed STAT3 phosphorylation and its upstage kinases, reduced expression of HIF1α, VEGF, VEGFR1, VEGFR2, and pro-inflammatory cytokines, and suppressed tumor volume and micro vessel density. The authors conclude that pentoxifylline may exert anti-tumor activity by inhibiting angiogenesis through the STAT3 pathway in B16F10 melanoma. | |
| Kamran & Gude, 2013 [46] | Intra-dermal mouse xenograft model was used to assess tumor volume and angiogenesis. IC50 7 mM after 2 h exposure with pentoxifylline. | A375 human melanoma cell line was treated with pentoxifylline and assessed for STAT3 signaling. | Following treatment of the mice with pentoxifylline, there was a significant decrease in the mean volume of the tumors and a reduction in tumor-induced angiogenesis. Pentoxifylline’s tumor growth and angiogenesis inhibition may involve the STAT3 signaling pathways. |
| Nidhyanandan et al., 2015 [47] | MS-275 and pentoxifylline were assessed in a murine Matrigel plug angiogenesis model and human breast cancer (MDA-MB-231) xenograft model. | A panel of cancer cell lines was treated with pentoxifylline and MS-275 and evaluated for cellular proliferation, cell cycle regulation, apoptosis, and anti-angiogenesis. | A combination of MS-275 and pentoxifylline significantly inhibited angiogenesis in the Matrigel plug angiogenesis assay. The combination therapy inhibited the expression of VEGF in a dose-dependent manner. |
| Çakmak et al., 2015 [35] | Sprague–Dawley rats were utilized to determine the effect of pentoxifylline on angiogenesis and bone healing. Radiographic, immunohistochemical methods and histological methods were utilized to evaluate the effect. | Pentoxifylline may improve angiogenesis and healing of segmental cortical bone defects of the radius in a rat model. | |
| Nathan et al., 2016 [36] | Various concentrations of pentoxifylline were tested at 50% epiboly stage (5.2 HPF) of zebrafish embryos and evaluated phenotypic changes and expression of adenosine receptors, HIF-1α, VEGFaa, VEGFr2, and RP-1a. | RBC staining demonstrated an absence of intersegmental vessels in embryos treated with pentoxifylline. Pentoxifylline-treated embryos developed abnormal vasculature. Additional results show inhibition of VEGFAA and adenosine receptions and new blood vessel formation following treatment with pentoxifylline. | |
| Bałan et al., 2017 [37] | Tumor cells were incubated with various concentrations of pentoxifylline before transplantation into mice. | The results of this study demonstrate that pentoxifylline had an inhibitory effect on tumor growth and volume and had a dose-dependent decrease in angiogenesis following transplantation. | |
| Niderla-Bielińska et al., 2018 [41] | Mouse embryo proepicardium was harvested and treated with pentoxifylline to assess the expression of angiogenic factors. Endothelial cell line C166 was derived from embryonic yolk sac treated with pentoxifylline to assess the direct effect on angiogenesis. | Pentoxifylline indirectly inhibits angiogenesis in mouse proepicardial explant cultures by decreasing Dll4 and Notch1 expression but has no significant effect on the C166 endothelial cell line. | |
| Yang et al., 2018 [38] | A mouse model was used to investigate pentoxifylline’s effect on postoperative intra-abdominal adhesion formation through angiogenesis and other physiological processes. Angiogenesis was assessed via immunohistology analysis of angiogenesis markers Ki67+/CD31+. | Pentoxifylline significantly suppressed angiogenesis during the peritoneal repair of the mice. The authors state that these findings are in line with additional studies on the inhibition of angiogenesis. | |
| Arsenyan et al., 2020 [42] | Matrigel (BD Biosciences) human umbilical vein endothelial cell tube formation model was used to investigate various compounds’ angiogenesis and MMP inhibition activity. | The study results show that pentoxifylline did not inhibit any of the studied matrix metalloproteinases. The authors presume that the lack of correlations between MMP and angiogenesis inhibition indicates that the compounds modulate angiogenesis via different mechanisms. | |
| Pedretti et al., 2020 [39] | Rat model of skin flap surgical procedure, then treated with subcutaneous pentoxifylline. VEGF and TGF-β1 levels were measured. | Pentoxifylline stimulated angiogenesis and reepithelization while reducing fibrogenesis. | |
| Seo et al., 2020 [40] | Rat model of radiation-induced osteoradionecrosis. Treated with pentoxifylline alone and with pentoxifylline and tocopherol. Angiogenesis effects were assessed. | Pentoxifylline and tocopherol work synergistically to promote angiogenesis, while pentoxifylline alone had a slight increase in proangiogenic factors PECAM, VEGF-A, and TNF-α. | |
| Seo et al., 2021 [43] | The effects of pentoxifylline on RAW 264.7 cells were analyzed with immunoprecipitation high-performance liquid chromatography to assess angiogenesis inhibition via the expression levels of VEGF-A, vWF, ET-1, CD31, MMP-10, and VCAM. | The expression levels of VEGF-A, vWF, ET-1, CD31, MMP-10, and VCAM showed a minimal change within 5%. The results of this study show that pentoxifylline has a weak angiogenic effect over 48 h. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khoury, W.; Trus, R.; Chen, X.; Baghaie, L.; Clark, M.; Szewczuk, M.R.; El-Diasty, M. Parsimonious Effect of Pentoxifylline on Angiogenesis: A Novel Pentoxifylline-Biased Adenosine G Protein-Coupled Receptor Signaling Platform. Cells 2023, 12, 1199. https://doi.org/10.3390/cells12081199
Khoury W, Trus R, Chen X, Baghaie L, Clark M, Szewczuk MR, El-Diasty M. Parsimonious Effect of Pentoxifylline on Angiogenesis: A Novel Pentoxifylline-Biased Adenosine G Protein-Coupled Receptor Signaling Platform. Cells. 2023; 12(8):1199. https://doi.org/10.3390/cells12081199
Chicago/Turabian StyleKhoury, William, Ryan Trus, Xingyu Chen, Leili Baghaie, Mira Clark, Myron R. Szewczuk, and Mohammad El-Diasty. 2023. "Parsimonious Effect of Pentoxifylline on Angiogenesis: A Novel Pentoxifylline-Biased Adenosine G Protein-Coupled Receptor Signaling Platform" Cells 12, no. 8: 1199. https://doi.org/10.3390/cells12081199