
Adhesion Molecules in Lung Inflammation from Repeated Glyphosate Exposures
 , and
, and 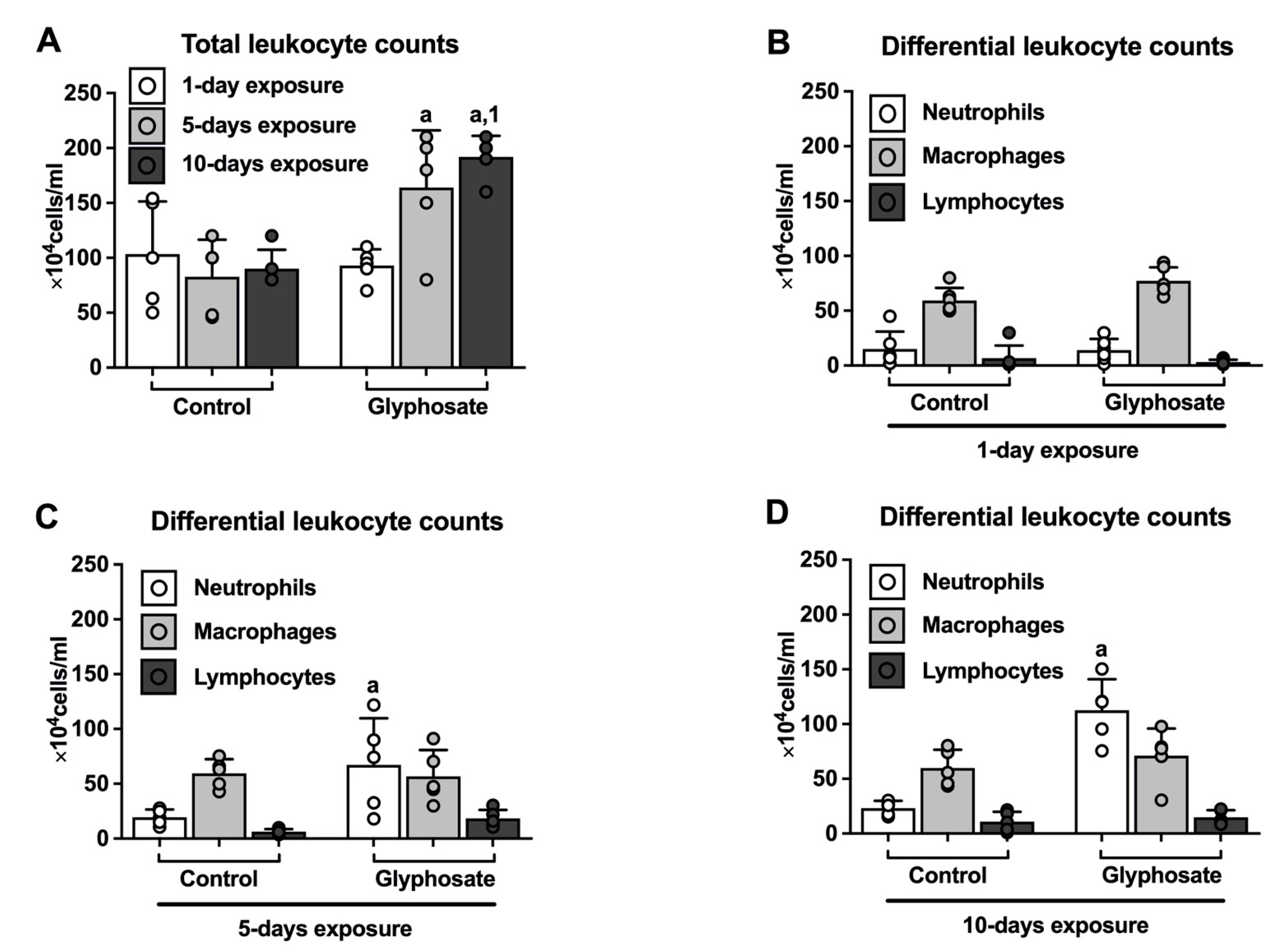{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice Exposure
2.2. Bronchoalveolar Lavage Fluid Collection and Processing
2.3. Lung Tissue Collection and Processing
2.4. Eosinophil Peroxidase Quantification
2.5. RNA Isolation and Real-Time PCR
2.6. Histology and Scoring for Inflammation
2.7. Immunohistochemistry and Analysis
2.8. Data Analysis and Statistics
3. Results
3.1. Leukocyte Counts in BAL
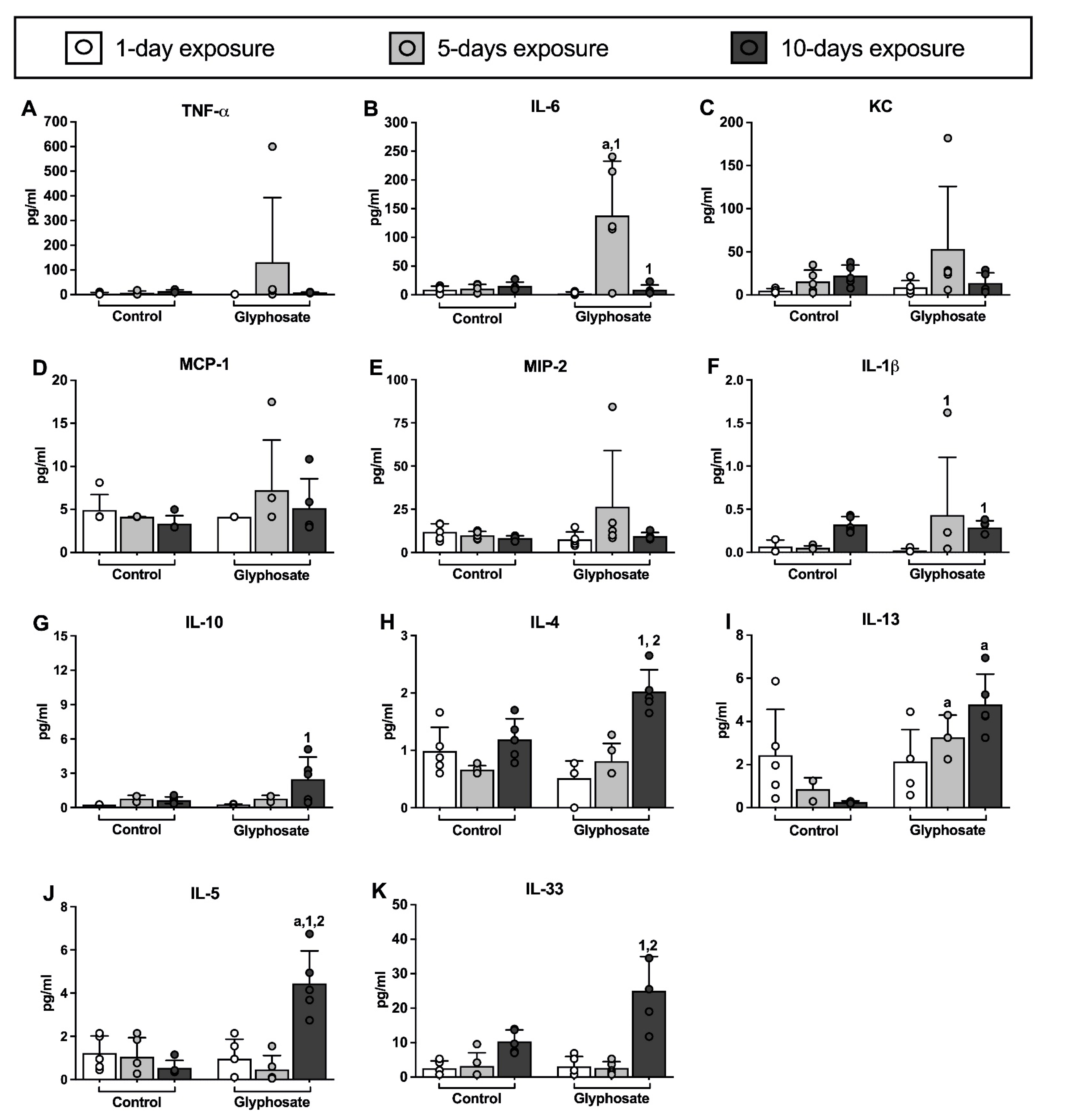
3.2. Cytokine Levels in BAL
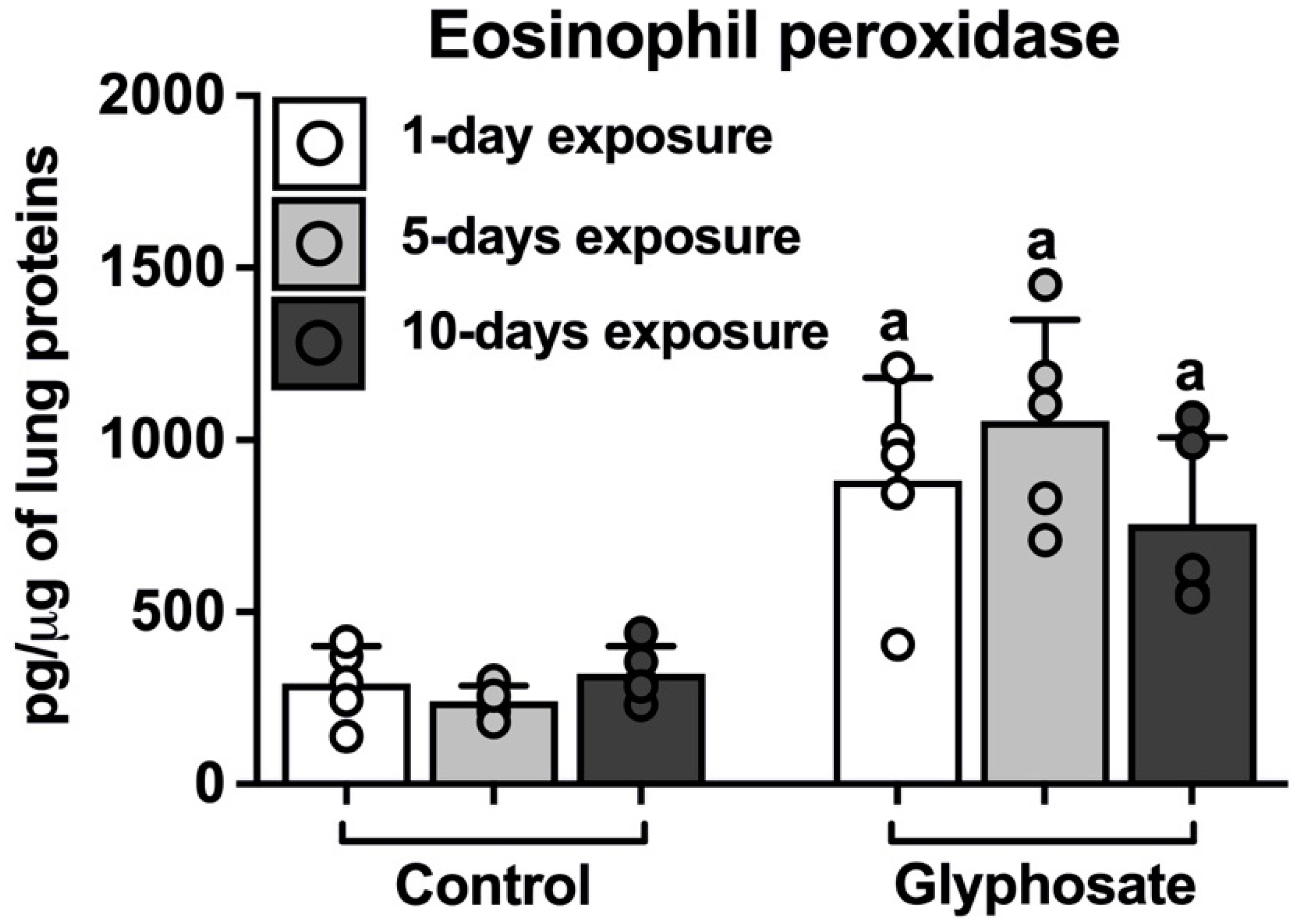
3.3. Eosinophil Peroxidase in Lungs
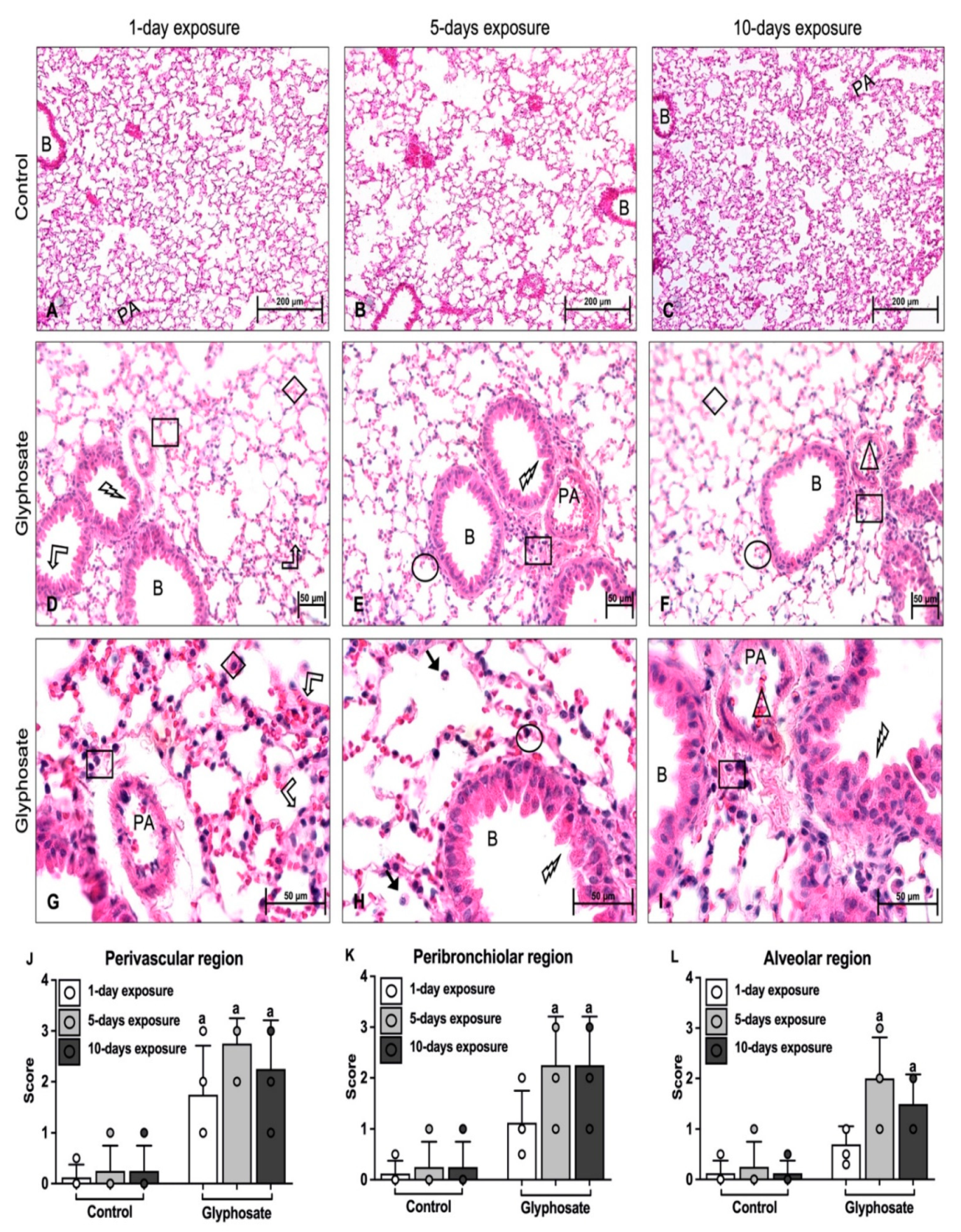
3.4. Lung Histology
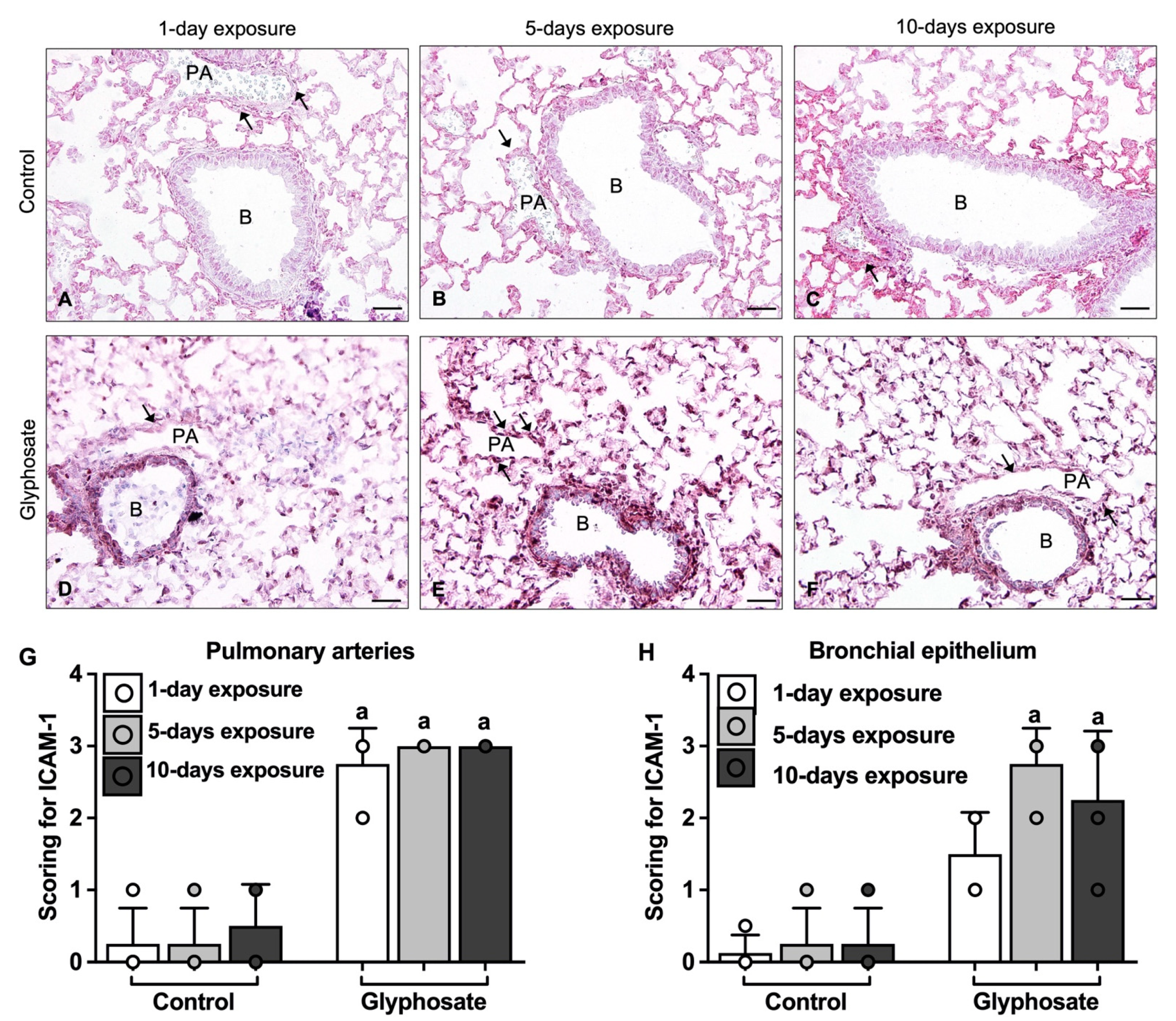
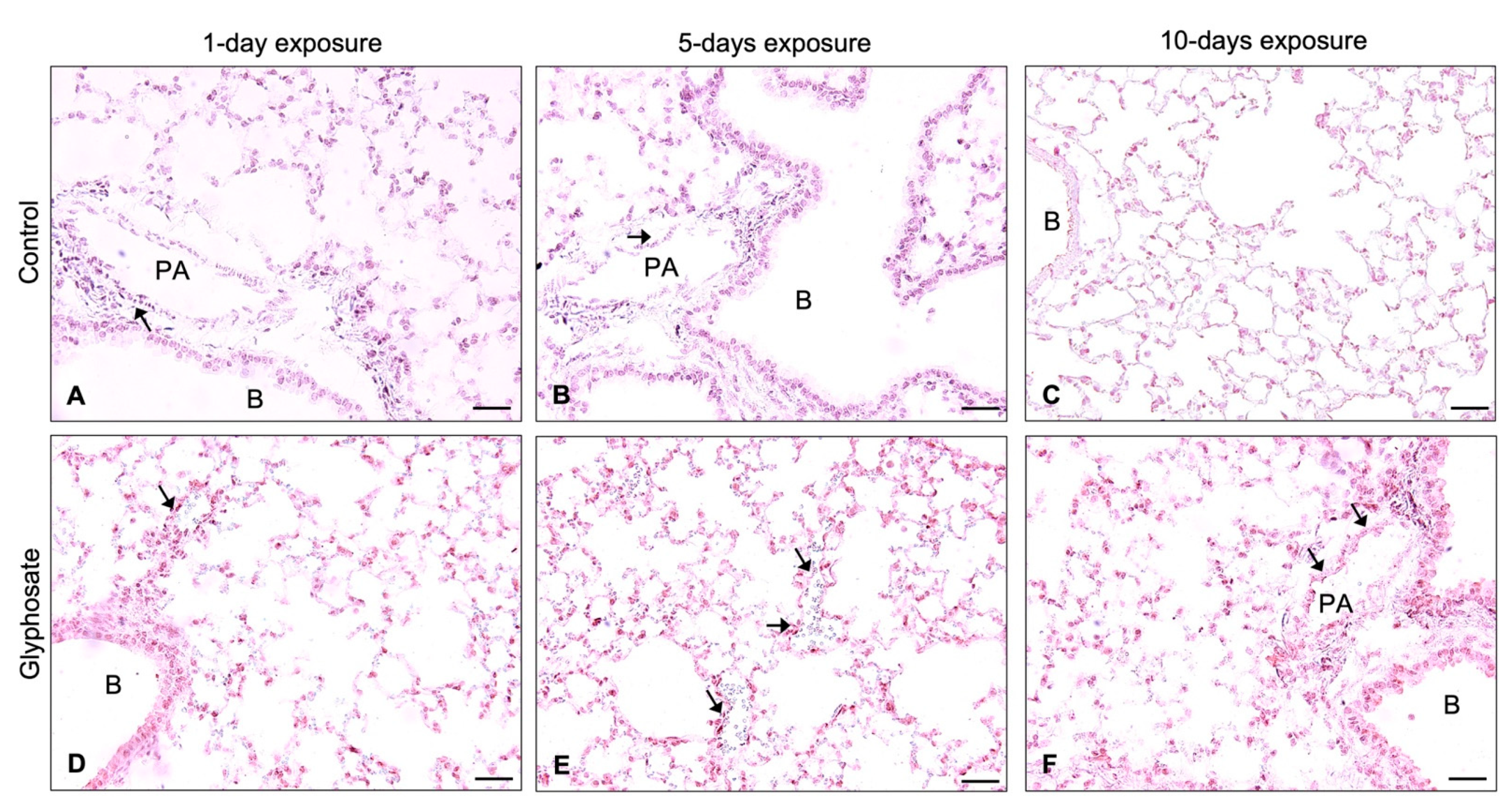
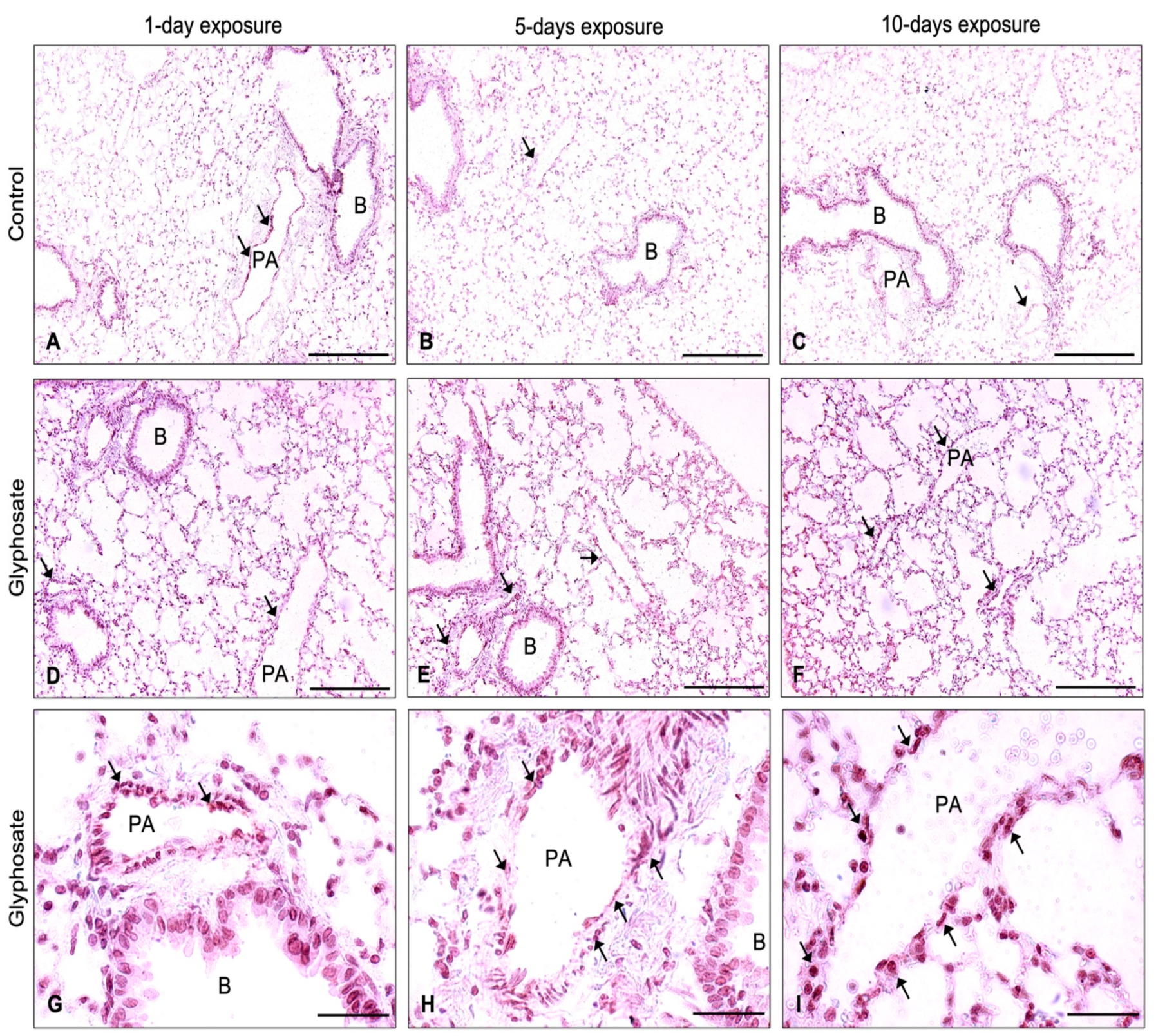
3.5. ICAM-1, VCAM-1, and vWF Staining
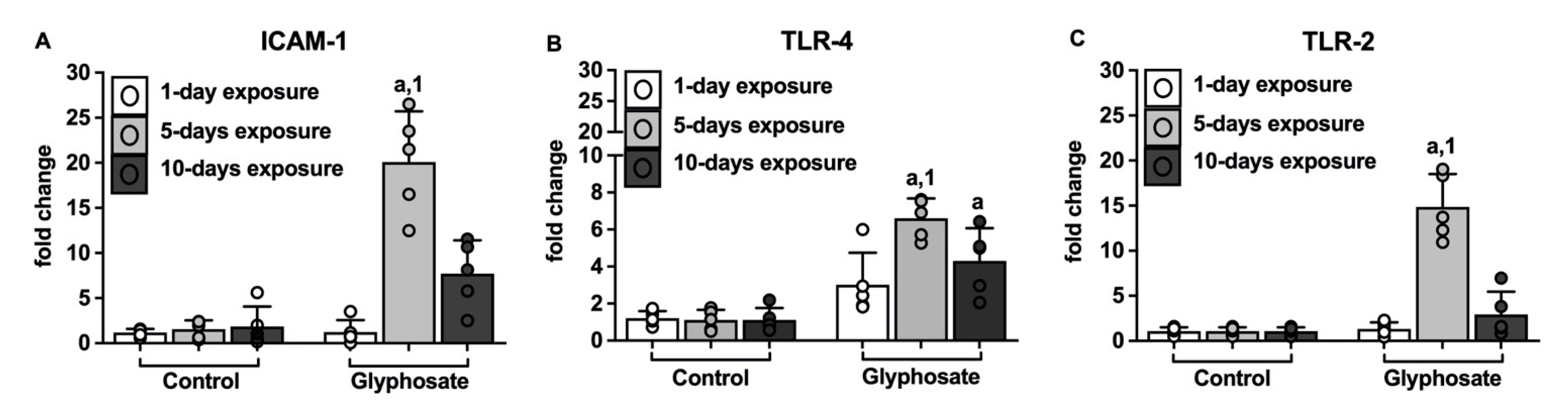
3.6. ICAM-1, TLR-4, and TLR-2 mRNA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benbrook, C.M. Trends in Glyphosate Herbicide Use in the United States and Globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Khodoun, M.; Kettleson, E.M.; McKnight, C.; Reponen, T.; Grinshpun, S.A.; Adhikari, A. Glyphosate-Rich Air Samples Induce IL-33, TSLP and Generate IL-13 Dependent Airway Inflammation. Toxicology 2014, 325, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Connolly, A.; Jones, K.; Galea, K.S.; Basinas, I.; Kenny, L.; McGowan, P.; Coggins, M. Exposure Assessment Using Human Biomonitoring for Glyphosate and Fluroxypyr Users in Amenity Horticulture. Int. J. Hyg. Environ. Health 2017, 220, 1064–1073. [Google Scholar] [CrossRef]
- Gillezeau, C.; van Gerwen, M.; Shaffer, R.M.; Rana, I.; Zhang, L.; Sheppard, L.; Taioli, E. The Evidence of Human Exposure to Glyphosate: A Review. Environ. Health 2019, 18, 2–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesnage, R.; Moesch, C.; Grand, R.L.G.; Lauthier, G.; de Vendômois, J.S.; Gress, S.; Séralini, G.-E. Glyphosate Exposure in a Farmer’s Family. J. Environ. Prot. Irvine Calif. 2012, 3, 1001–1003. [Google Scholar] [CrossRef] [Green Version]
- Hoppin, J.A.; Umbach, D.M.; London, S.J.; Henneberger, P.K.; Kullman, G.J.; Alavanja, M.C.R.; Sandler, D.P. Pesticides and Atopic and Nonatopic Asthma among Farm Women in the Agricultural Health Study. Am. J. Respir. Crit. Care Med. 2008, 177, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppin, J.A.; Umbach, D.M.; London, S.J.; Henneberger, P.K.; Kullman, G.J.; Coble, J.; Alavanja, M.C.R.; Beane Freeman, L.E.; Sandler, D.P. Pesticide Use and Adult-Onset Asthma among Male Farmers in the Agricultural Health Study. Eur. Respir. J. 2009, 34, 1296–1303. [Google Scholar] [CrossRef]
- Hoppin, J.A.; Umbach, D.M.; Long, S.; London, S.J.; Henneberger, P.K.; Blair, A.; Alavanja, M.; Freeman, L.E.B.; Sandler, D.P. Pesticides Are Associated with Allergic and Non-Allergic Wheeze among Male Farmers. Environ. Health Perspect. 2017, 125, 535–543. [Google Scholar] [CrossRef]
- Slager, R.E.; Poole, J.A.; LeVan, T.D.; Sandler, D.P.; Alavanja, M.C.R.; Hoppin, J.A. Rhinitis Associated with Pesticide Exposure among Commercial Pesticide Applicators in the Agricultural Health Study. Occup. Environ. Med. 2009, 66, 718–724. [Google Scholar] [CrossRef]
- Slager, R.E.; Simpson, S.L.; Levan, T.D.; Poole, J.A.; Sandler, D.P.; Hoppin, J.A. Rhinitis Associated with Pesticide Use among Private Pesticide Applicators in the Agricultural Health Study. J. Toxicol. Environ. Health—Part A Curr. Issues 2010, 73, 1382–1393. [Google Scholar] [CrossRef] [Green Version]
- Henneberger, P.K.; Liang, X.; London, S.J.; Umbach, D.M.; Sandler, D.P.; Hoppin, J.A. Exacerbation of Symptoms in Agricultural Pesticide Applicators with Asthma. Int. Arch. Occup. Environ. Health 2014, 87, 423–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilewski, J.M.; Albelda, S.M. Adhesion Molecules in the Lung: An Overview. Am. Rev. Respir. Dis. 1993, 148, S31–S37. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, J.; Schaberg, T. Adhesion Molecules in Lung Diseases. Lung 1994, 172, 189–213. [Google Scholar] [CrossRef]
- Osborn, L. Leukocyte Adhesion to Endothelium in Inflammation. Cell 1990, 62, 3–6. [Google Scholar] [CrossRef]
- Montefort, S.; Feather, I.H.; Wilson, S.J.; Haskard, D.O.; Lee, T.H.; Holgate, S.T.; Howarth, P.H. The Expression of Leukocyte-Endothelial Adhesion Molecules Is Increased in Perennial Allergic Rhinitis. Am. J. Respir. Cell Mol. Biol. 1992, 7, 393–398. [Google Scholar] [CrossRef]
- Montefort, S.; Lai, C.K.W.; Kapahi, P.; Leung, J.; Lai, K.N.; Chan, H.S.; Haskard, D.O.; Howarth, P.H.; Holgate, S.T. Circulating Adhesion Molecules in Asthma. Am. J. Respir. Crit. Care Med. 1994, 149, 1149–1152. [Google Scholar] [CrossRef] [PubMed]
- Beck-Schimmer, B.; Madjdjpour, C.; Kneller, S.; Ziegler, U.; Pasch, T.; Wüthrich, R.P.; Ward, R.P.; Schimmer, R.C. Role of Alveolar Epithelial ICAM-1 in Lipopolysaccharide-Induced Lung Inflammation. Eur. Respir. J. 2002, 19, 1142–1150. [Google Scholar] [CrossRef]
- Beck-Schimmer, B.; Schimmer, R.C.; Warner, R.L.; Schmal, H.; Nordblom, G.; Flory, C.M.; Lesch, M.E.; Friedl, H.P.; Schrier, D.J.; Ward, P.A. Expression of Lung Vascular and Airway ICAM-1 after Exposure to Bacterial Lipopolysaccharide. Am. J. Respir. Cell Mol. Biol. 1997, 17, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Springer, T.A. Adhesion Receptors of the Immune System. Nature 1990, 346, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Vadas, M.; Gamble, J.; Smith, W. Regulation of Myeloid Blood Cell-Endothelial Interaction by Cytokines. In Adhesion. Its Role in Inflammatory Disease; Harlan, J.M., Liu, D.Y., Eds.; W.H. Freeman: New York, NY, USA, 1992; pp. 65–81. [Google Scholar]
- Pandher, U.; Kirychuk, S.; Schneberger, D.; Thompson, B.; Aulakh, G.; Sethi, R.S.; Singh, B. Pulmonary Inflammatory Response from Co-Exposure to LPS and Glyphosate. Environ. Toxicol. Pharmacol. 2021, 86, 103651. [Google Scholar] [CrossRef] [PubMed]
- Pandher, U.; Kirychuk, S.; Schneberger, D.; Thompson, B.; Aulakh, G.; Sethi, R.S.; Singh, B. Lung Inflammation from Repeated Exposure to LPS and Glyphosate. Cell Tissue Res. 2021, 386, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Quinlan, W.M.; Doyle, N.A.; Graham, L.; Sligh, J.E.; Takei, F.; Beaudet, A.L.; Doerschuk, C.M. The roles of CD11/CD18 and ICAM-1 in acute Pseudomonas aeruginosa-induced pneumonia in mice. J. Immunol. 1996, 157, 5016–5021. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Sano, H.; Nishimura, T.; Yoshida, S.; Iwamoto, I. Role of vascular cell adhesion molecule 1/very later activation antigen 4 and intercellular adhesion molecule 1/lymphocyte function-associated antigen 1 interactions in antigen-induced eosinophil and T cell recruitment into the tissue. J. Exp. Med. 1994, 179, 1145–1154. [Google Scholar] [CrossRef]
- Lee, J.; Sohn, J.; Ryu, S.Y.; Hong, C.; Moon, K.D.; Park, J. A novel human anti-VCAM-1 monoclonal antibody ameliorates airway inflammation and remodelling. J. Cell. Mol. Med. 2013, 17, 1271–1281. [Google Scholar] [CrossRef]
- Lötvall, J.; Pullerits, T. Treating Asthma with Anti-IgE or Anti-IL5. Curr. Pharm. Des 1999, 5, 757–770. [Google Scholar] [CrossRef]
- Zhu, Z.; Lee, C.G.; Zheng, T.; Chupp, G.; Wang, J.; Homer, R.J.; Noble, P.W.; Hamid, Q.; Elias, J.A. Airway Inflammation and Remodeling in Asthma. Am. J. Respir. Crit. Care Med. 2001, 164, S67–S70. [Google Scholar] [CrossRef]
- Booth, B.W.; Adler, K.B.; Bonner, J.C.; Tournier, F.; Martin, L.D. Interleukin-13 Induces Proliferation of Human Airway Epithelial Cells In Vitro via a Mechanism Mediated by Transforming Growth Factor-α. Am. J. Respir. Cell Mol. Biol. 2001, 25, 739–743. [Google Scholar] [CrossRef]
- Gharaee-Kermani, M.; Phan, S.H. Lung Interleukin-5 Expression in Murine Bleomycin-Induced Pulmonary Fibrosis. Am. J. Respir. Cell Mol. Biol. 1997, 16, 438–447. [Google Scholar] [CrossRef]
- Kim, H.Y.; Chang, Y.J.; Subramanian, S.; Lee, H.H.; Albacker, L.A.; Matangkasombut, P.; Savage, P.B.; McKenzie, A.N.J.; Smith, D.E.; Rottman, J.B.; et al. Innate Lymphoid Cells Responding to IL-33 Mediate Airway Hyperreactivity Independently of Adaptive Immunity. J. Allergy Clin. Immunol. 2012, 129, 216–227.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabryelska, A.; Kuna, P.; Antczak, A.; Białasiewicz, P.; Panek, M. IL-33 Mediated Inflammation in Chronic Respiratory Diseases—Understanding the Role of the Member of IL-1 Superfamily. Front. Immunol. 2019, 10, 692. [Google Scholar] [CrossRef] [PubMed]
- Heyen, L.; Müller, U.; Siegemund, S.; Schulze, B.; Protschka, M.; Alber, G.; Piehler, D. Lung Epithelium Is the Major Source of IL-33 and Is Regulated by IL-33-Dependent and IL-33-Independent Mechanisms in Pulmonary Cryptococcosis. Pathog. Dis. 2016, 74, ftw086. [Google Scholar] [CrossRef] [Green Version]
- Bonfield, T.L.; Konstan, M.W.; Burfeind, P.; Panuska, J.R.; Hilliard, J.B.; Berger, M. Normal Bronchial Epithelial Cells Constitutively Produce the Anti-Inflammatory Cytokine Interleukin-10, Which Is Downregulated in Cystic Fibrosis. Am. J. Respir. Cell Mol. Biol. 1995, 13, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.E. IL-33: A Tissue Derived Cytokine Pathway Involved in Allergic Inflammation and Asthma. Clin. Exp. Allergy 2010, 40, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y.; Pitman, N.I.; McInnes, I.B. Disease-Associated Functions of IL-33: The New Kid in the IL-1 Family. Nat. Rev. Immunol. 2010, 10, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Oboki, K.; Ohno, T.; Kajiwara, N.; Arae, K.; Morita, H.; Ishii, A.; Nambu, A.; Abe, T.; Kiyonari, H.; Matsumoto, K.; et al. IL-33 Is a Crucial Amplifier of Innate Rather than Acquired Immunity. Proc. Natl. Acad. Sci. USA 2010, 107, 18581–18586. [Google Scholar] [CrossRef] [Green Version]
- Cleary, S.J.; Hobbs, C.; Amison, R.T.; Arnold, S.; O’Shaughnessy, B.G.; Lefrançais, E.; Mallavia, B.; Looney, M.R.; Page, C.P.; Pitchford, S.C. LPS-Induced Lung Platelet Recruitment Occurs Independently from Neutrophils, PSGL-1, and P-Selectin. Am. J. Respir. Cell Mol. 2019, 61, 232–243. [Google Scholar] [CrossRef]
- Schwartz, D.A.; Thorne, P.S.; Jagielo, P.J.; White, G.E.; Bleuer, S.A.; Frees, K.L. Endotoxin Responsiveness and Grain Dust-Induced Inflammation in the Lower Respiratory Tract. Am. J. Physiol.-Lung Cell. Mol. Physiol. 1994, 267, L609–L617. [Google Scholar] [CrossRef]
- Senthilselvan, A.; Chénard, L.; Grover, V.; Kirychuk, S.; Hagel, L.; Ulmer, K.; Hurst, T.S.; Dosman, J.A. Excess Longitudinal Decline in Lung Function in Grain Farmers. J. Agromed. 2010, 15, 157–165. [Google Scholar] [CrossRef]
- Von Essen, S. The Role of Endotoxin in Grain Dust Exposure and Airway Obstruction. Curr. Opin. Pulm. Med. 1997, 3, 198–202. [Google Scholar] [CrossRef]
- Joosten, L.; Abdollahi-Roodsaz, S.; Dinarello, C.A.; O’Neill, L.; Netea, M.G. Toll-like Receptors and Chronic Inflammation in Rheumatic Diseases: New Developments. Nat. Rev. Rheumatol. 2016, 12, 344–357. [Google Scholar] [CrossRef]
- Janardhan, K.S.; McIsaac, M.; Fowlie, J.; Shrivastav, A.; Caldwell, S.; Sharma, R.K.; Singh, B. Toll like Receptor-4 Expression in Lipopolysaccharide Induced Lung Inflammation. Histol. Histopathol. 2006, 21, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Di Candia, L.; Gomez, E.; Venereau, E.; Chachi, L.; Kaur, D.; Bianchi, M.E.; Challiss, R.A.J.; Brightling, C.E.; Saunders, R.M. HMGB1 Is Upregulated in the Airways in Asthma and Potentiates Airway Smooth Muscle Contraction via TLR4. J. Allergy Clin. Immunol. 2017, 140, 584–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haczku, A. Protective Role of the Lung Collectins Surfactant Protein A and Surfactant Protein D in Airway Inflammation. J. Allergy Clin. Immunol. 2008, 122, 861–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, K.; Akira, S. TLR Signaling Pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef]
- Frey, A.; Lunding, L.P.; Ehlers, J.C.; Weckmann, M.; Zissler, U.M.; Wegmann, M. More Than Just a Barrier: The Immune Functions of the Airway Epithelium in Asthma Pathogenesis. Front. Immunol. 2020, 11, 761. [Google Scholar] [CrossRef]
- Post, S.; Nawijn, M.C.; Jonker, M.R.; Kliphuis, N.; Van Den Berge, M.; Van Oosterhout, A.J.M.; Heijink, I.H. House Dust Mite-Induced Calcium Signaling Instigates Epithelial Barrier Dysfunction and CCL20 Production. Allergy Eur. J. Allergy Clin. Immunol. 2013, 68, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Soyka, M.B.; Wawrzyniak, P.; Eiwegger, T.; Holzmann, D.; Treis, A.; Wanke, K.; Kast, J.I.; Akdis, C.A. Defective Epithelial Barrier in Chronic Rhinosinusitis: The Regulation of Tight Junctions by IFN-γ and IL-4. J. Allergy Clin. Immunol. 2012, 130, 1087–1096.e10. [Google Scholar] [CrossRef]
- Chakraborty, S.; Mukherjee, S.; Roychoudhury, S.; Siddique, S.; Lahiri, T.; Ray, M.R. Chronic Exposures to Cholinesterase-inhibiting Pesticides Adversely Affect Respiratory Health of Agricultural Workers in India. J. Occup. Health 2009, 51, 488–497. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Beach, J.; Martin, J.W.; Senthilselvan, A. Occupational Pesticide Exposures and Respiratory Health. Int. J. Environ. Res. Public Health 2013, 10, 6442–6471. [Google Scholar] [CrossRef]
- Hernández, A.F.; Casado, I.; Pena, G.; Gil, F.; Villanueva, E.; Pla, A. Low Level of Exposure to Pesticides Leads to Lung Dysfunction in Occupationally Exposed Subjects. Inhal. Toxicol. 2008, 20, 839–849. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandher, U.; Kirychuk, S.; Schneberger, D.; Thompson, B.; Aulakh, G.; Sethi, R.S.; Singh, B. Adhesion Molecules in Lung Inflammation from Repeated Glyphosate Exposures. Int. J. Environ. Res. Public Health 2023, 20, 5484. https://doi.org/10.3390/ijerph20085484
Pandher U, Kirychuk S, Schneberger D, Thompson B, Aulakh G, Sethi RS, Singh B. Adhesion Molecules in Lung Inflammation from Repeated Glyphosate Exposures. International Journal of Environmental Research and Public Health. 2023; 20(8):5484. https://doi.org/10.3390/ijerph20085484
Chicago/Turabian StylePandher, Upkardeep, Shelley Kirychuk, David Schneberger, Brooke Thompson, Gurpreet Aulakh, R. S. Sethi, and Baljit Singh. 2023. "Adhesion Molecules in Lung Inflammation from Repeated Glyphosate Exposures" International Journal of Environmental Research and Public Health 20, no. 8: 5484. https://doi.org/10.3390/ijerph20085484